



Survey of European mites from the suborder Uropodina: II. Morphology, geographical distribution, biology, and ecology of Trematurella elegans (Kramer, 1882)
Błoszyk, Jerzy1 ; Adamski, Zbigniew2 and Napierała, Agnieszka 3
1✉ Department of General Zoology, Faculty of Biology, Adam Mickiewicz University, ul. Umultowska 89;61-614 Poznań, Poland. Natural History Collections, Faculty of Biology, Adam Mickiewicz University, ul. Umultowska 89; 61-614 Poznań, Poland.
2Electron and Confocal Microscope Laboratory/4 Department of Animal Physiology and Development, Faculty of Biology, Adam Mickiewicz University, ul. Umultowska 89; 61-614 Poznań, Poland.
3Department of General Zoology, Faculty of Biology, Adam Mickiewicz University, ul. Umultowska 89;61-614 Poznań, Poland.
2018 - Volume: 58 Issue: 3 pages: 683-709
https://doi.org/10.24349/acarologia/20184265ZooBank LSID: 75AAB6E7-7BD2-49DA-8948-49E8960EF327
Keywords
Abstract
This article is dedicated to the memory of an outstanding acarologist, our colleague and teacher, Dr. Françoise Athias Binche
It may seem that European Uropodina mites have been already described so thoroughly that it is virtually impossible to provide any new information about mites from this suborder. Since these mites are quite large and very characteristic, many species had been described in the late 1800’s and the early 1900’s, but those descriptions are not very precise (see e.g. Müller 1776; Hermann 1804; Haller 1881; Koch 1835, 1839, 1941, 1847; Berlese 1888, 1913, 1917; Michael 1894; Halbert 1915, 1923). At that time the researchers focused mostly on the taxonomy, and they described the species which were new for science and paid less attention to the biology, ecology, and geographical distribution of the species. For this reason the idea of more detail-oriented research into the most abundant European species appeared later. The first publication from a series was published by Błoszyk and Athias-Binche (1998), who described Polyaspinus cylindricus Berlese, 1916, a species which has very interesting geographical distribution and ecology.
Nowadays, professor Jerzy Błoszyk, with his co-workers, decided to go back to the idea of a renewed elaboration of the morphology, biology, ecology, and zoogeography of the most interesting European Uropodina species. The current article focuses on Trematurella elegans, which was described by Kramer 136 years ago, in 1882, in Germany.
The re-descriptions of selected European Uropodina will be prepared on the basis of the information available in the existing literature published so far, as well as unpublished information, gathered during over 50-year-long research into this group of mites. The next reason is that many of those species were described only in Italian or German, which made the descriptions inaccessible for many readers. Moreover, the authors of the current article also want to make their own contribution towards the research by the use of modern research techniques, like scanning electron microscopy (SEM) for the morphological analysis, and the GIS systems to establish the distribution of Uropodina (Błoszyk et al. 2013).
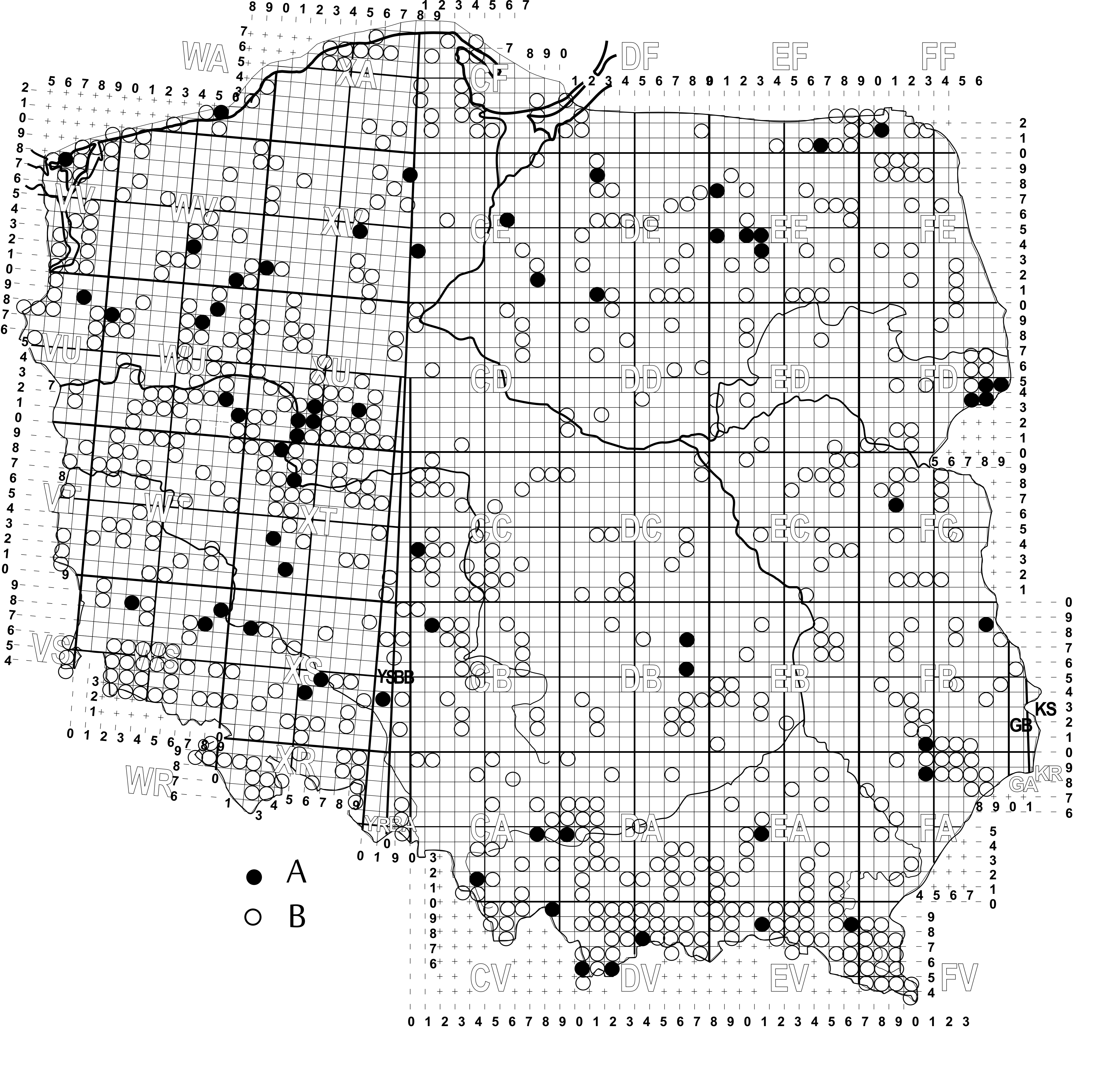
The study is based on 21,741quantitative and qualitive samples, which were collected in Poland between 1961 and 2017. The samples were collected in areas with different altitude up to over 2,400 m above sea level. The analysis of the geographical distribution of T. elegans in Europe (Figure 14) was carried out on the basis of data available in the literature and data collected through examination of soil samples from Poland and different European countries: Spain (30 samples), France (50 samples), Belgium (40 samples), The Netherlands (100 samples), Germany (50 samples), Denmark (100 samples), Sweden (30 samples), Norway (500 samples), Czech Republic (150 samples), Slovakia (170 samples), and Ukraine (30 samples).
The samples were collected in different seasons from 35 types of habitats, such as open habitats, forests, and merocenoses (Table 1). The samples consisted of soil were collected either quantitatively with a corer (30-90 cm2) to the depth of 10 cm or qualitatively (with a sieve) from the forest floor, as well as unsieved samples from open habitats. The material from dead wood (such as rotten trunks, logs, stumps) was collected with a plastic container with a volume of 0.5–0.8 l. Mites were extracted using Tullgren funnels for 4–6 days (depending on the level of moisture) just after the material had been collected. The extracted mites were preserved in 75% ethyl alcohol. The specimens were identified using the morphological criteria from the original descriptions and later accounts (Kramer 1882; Hirschmann and Zirgiebl-Nicol 1961; Błoszyk 1983, 1999; Karg 1989; Mašan 2001).
To analyze the preferences towards specific habitat type, altitude, and phenology, over 7,000 qualitative samples were examined. The number of samples slightly varied between the tested phenomena, since the complete set of data necessary for the analysis was not always present. Therefore, 7,476, 7,190 and 7,506 samples were used to analyze habitat preferences, phenology, altitude preferences, respectively. The map of distribution of T. elegans in Poland was drawn on the basis of all collected samples (21,741) on the UTM grid (10×10 km).
To visualize the fine details of morphology of the mites, scanning electron microscopy (SEM) techniques were used. The specimens were air-dried, mounted on a pin stubs with a double-sided sticky tape, coated with gold in a sputter coater and observed in a Zeiss Evo 40 Scanning Electron Microscope (Carl Zeiss SMTLtd., Cambridge, UK) at 16.5 kV.
The analysis of frequency of T. elegans in Uropodina communities was based on the index of frequency. The following classes were used (Błoszyk 1999): Frequency: F5, euconstants (>50%), F4, constants (30.1–50%), F3, subconstants (15.1–30.0%), F2, accessory species (5.0–15.0%), and F1, accidents (<5%).
Trematurella Trägårdh, 1942: 111.
Type species Trematurella stylifera Trägårdh, 1942: 111, by monotypy.
Diagnosis — Medium size mites, highly sclerotized, with characteristic sculpture. The idiosoma is oval and flat at the dorsal part. The unique characteristics of this species: the large dorsal shield is surrounded by a rather narrow marginal shield. The dorsal setae are serrated, very long and massive, of the same width at the whole length. The female genital shield is large, wide at the base and becomes more and more narrow towards the front (the shape resembles the foot of the iron), with a long narrow appendix at the front, intensively sculptured. The male genital shield is oval, smooth and has no paragenital setae. They have five pairs of sternal setae, two pairs of adanal setae (Ad1-Ad2), and single postanal seta (Pa). There are cavities for the leg hiding, the last ones, hiding the fourth pair of legs, are located across and do not overlap with the opisthosoma. At the first pair of tarsus there is a single claw. The first pair of hypostomal setae (h1) is needle-shaped and smooth, the other ones (h2-h4) are feather-like. Also the setae at the trochanters of palps are feather-like.
Uropoda elegans Kramer, 1882: 406-407
Urodinychus elegans var. gallica – Berlese, 1917: 144-145.
Trematurella stylifera – Trӓgårdh, 1942: 109-112; Trӓgårdh – 1945: 1-10; Hirschmann & Zirngiebl-Nicol, 1961: 10; Wiśniewski, 1979: 37.
Trematurella elegans – Błoszyk, 1984: 70; Wiśniewski & Hirschmann, 1991: 174; Błoszyk, 1999: 136-137; Błoszyk & Krysiak, 2000: 117, 119; Błoszyk et al., 2002a 25, 29, 31; Błoszyk et al., 2002b 63, 67, 69-70; Błoszyk et al., 2004: 1507; Błoszyk & Napierała, 2004: 287-288; Błoszyk et al., 2006: 28, 32-33, 35; Napierała et al., 2006: 157; Napierała, 2008: 61, 90, 109, 111, 116, 149; Napierała et al., 2009: 291, 295, 299, 302; Kontschán, 2013: 103, 122; Napierała & Błoszyk, 2013: 166, 169, 175; 176; Błoszyk et al., 2015: 7, 10.
Trichouropoda elegans – Kontschán, 2002: 346; Kontschán, 2003: 186; Fenda & Ciceková, 2007: 30-31; Kontschán, 2008: 6; Kaczmarek et al., 2011: 33; Kazemi & Kontschán, 2014: 12; Huhta, 2016: 132.
Notes — Berlese (1917) in his work described this species as Urodinychus elegans Kram. var. gallicus Berl. n. var. This variation was also noted by Wiśniewski and Hirschmann (1991) in their catalogue. However, from the taxonomic point of view, it seems that there is no reason to distinguish this variation. With no doubt there is only one such species in Europe, and the differences stem from the range of geographical variability of the species.
The size of the body (original units used by the authors):
Kramer 1882: length 700 μm, width 560 μm;
Berlese 1917: length 650 μm, width 420 μm;
Błoszyk 1983, 1999: female 650 – 730 μm x 415 – 480 μm; male 650 – 700 μm x 400 – 450 μm;
Karg 1989: female length 670 μm; male length 640 μm;
Wiśniewski & Hirschmann 1993: female 670 x 470 μm; male 640 x 420 μm; deutonymphs 660 – 725 x 420 – 485 μm; deutonymphs (W) 535 – 565 x 335 – 345 μm; protonymphs 570 x 310 μm; larvae 330 x 220 μm;
Mašan 2001: length 535 – 625 μm;
The size of the body measured in this study:
Female (N=30) length 608 – 730 μm, width 401 – 480 μm, average: length 656 ± 44,1 SD μm, width 425 ± 25,9 SD μm
Male (N=27) length 596 – 700 μm, width 383 – 450 μm, average: length 630 ± 29,4 SD μm, width 416 ± 25,1. SD μm
Deutonymph (N=32) length 511 – 654 μm, width 332 – 479 μm, average: length 587 ± 36,2 SD μm, width 391 ± 35,2 SD μm
Protonymph (N=33) length 405 – 580 μm, width 246 – 356 μm, average: length 480 ± 42,8 SD μm, width 294 ± 30,9 SD μm
Larva (N=17) length 330 – 368 μm, width 189 – 239 μm, average: length 340 ± 13,4 SD μm, width 207 ± 17,0 SD μm
Female.
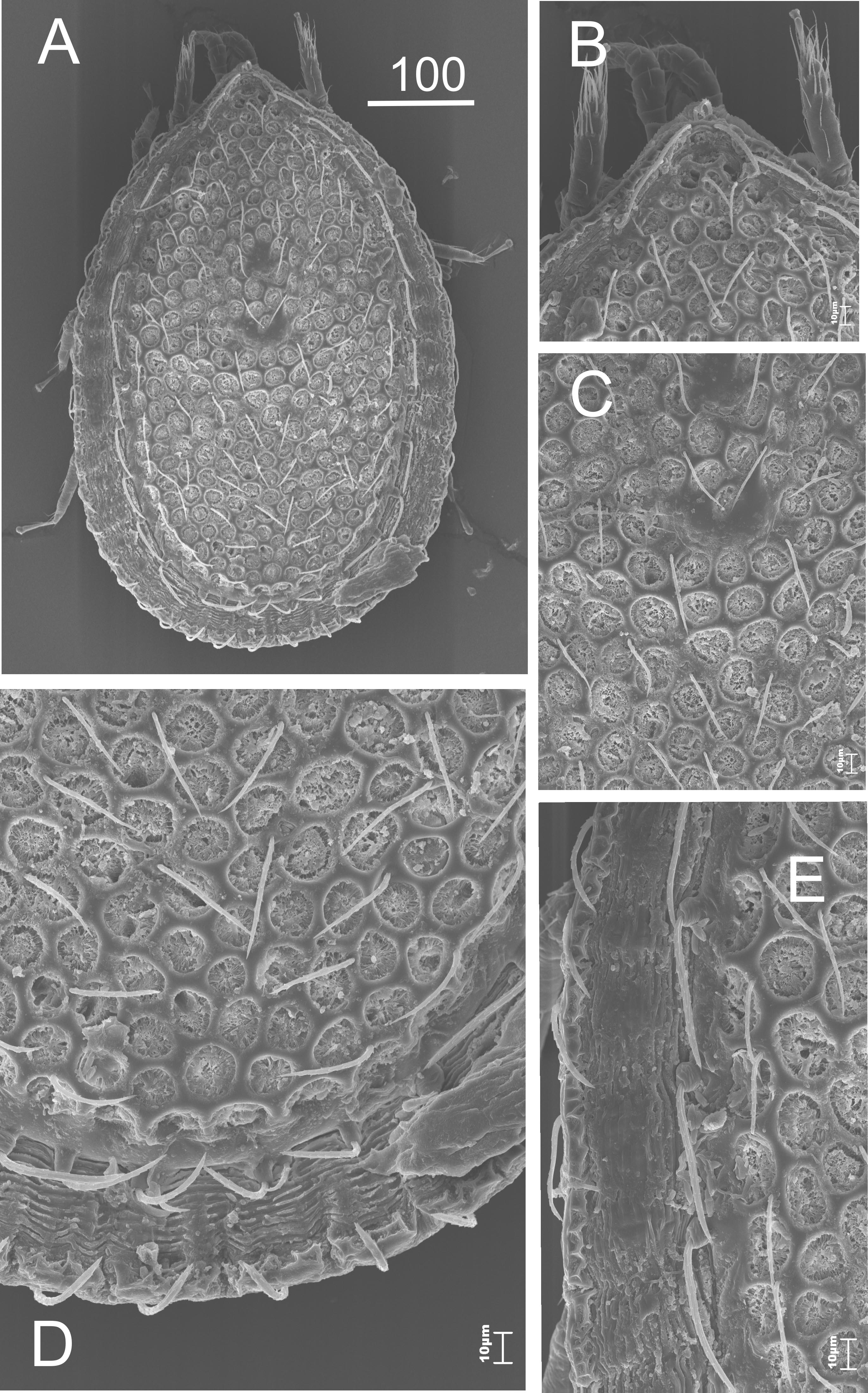
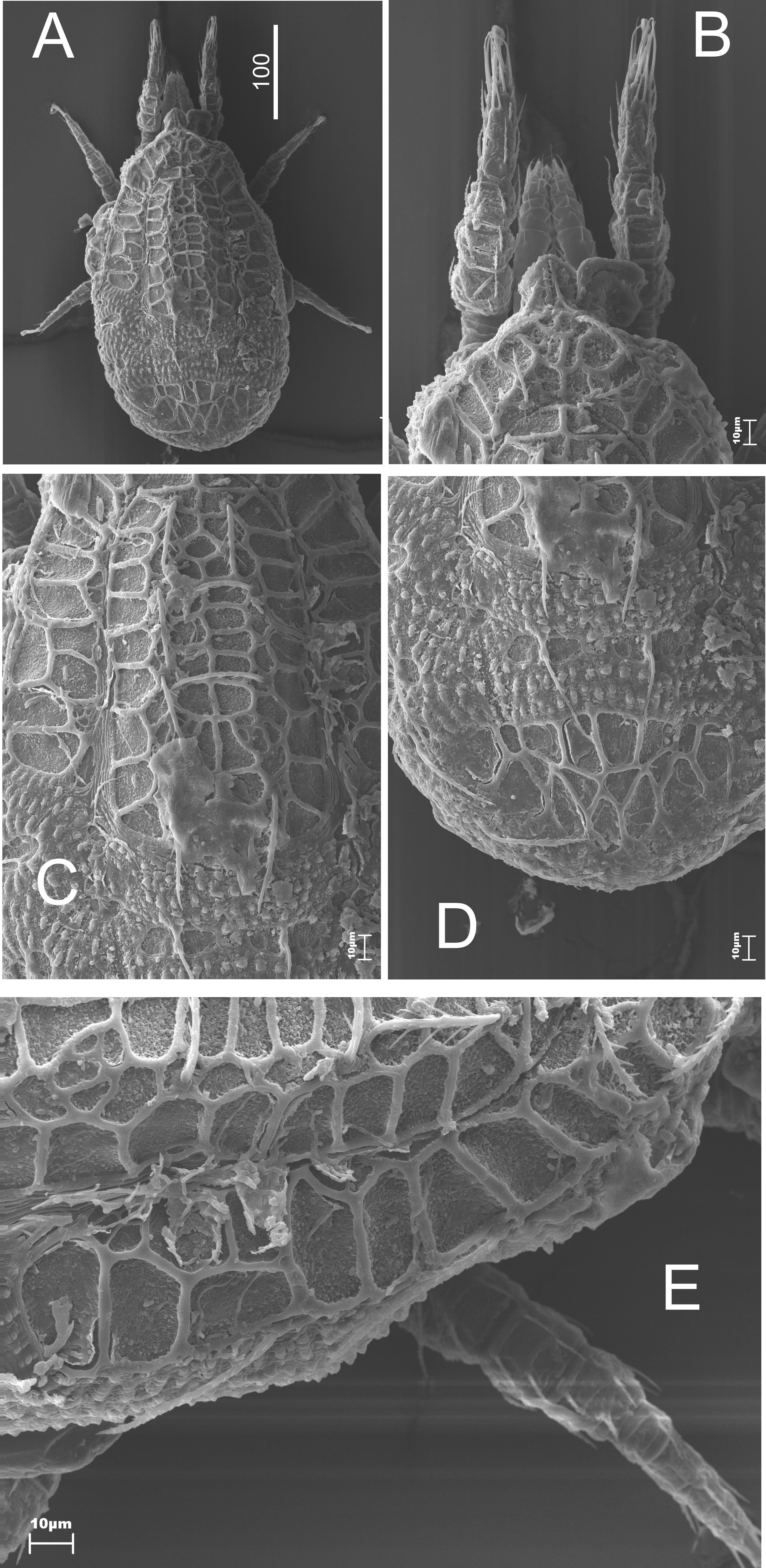
The dorsal side (Figures 1 and 2A, B). An oval-shaped idiosoma with a triangle-like vertex at the front edge (Figures 1A and 1B). The dorsal side is flat, highly sculptured. The narrow decorated marginal shield surrounds the big dorsal shield. Both shields fused at the anterior of the body. The whole surface of the dorsal shield is sculptured (Figures 1A and 1C-E). This sculpture is made of polygonal relief of chitin (Figure 2A). The dorsal setae are long, linear, serrated. The number of the setae can be different on each side of a specimen (cosmotrichy).

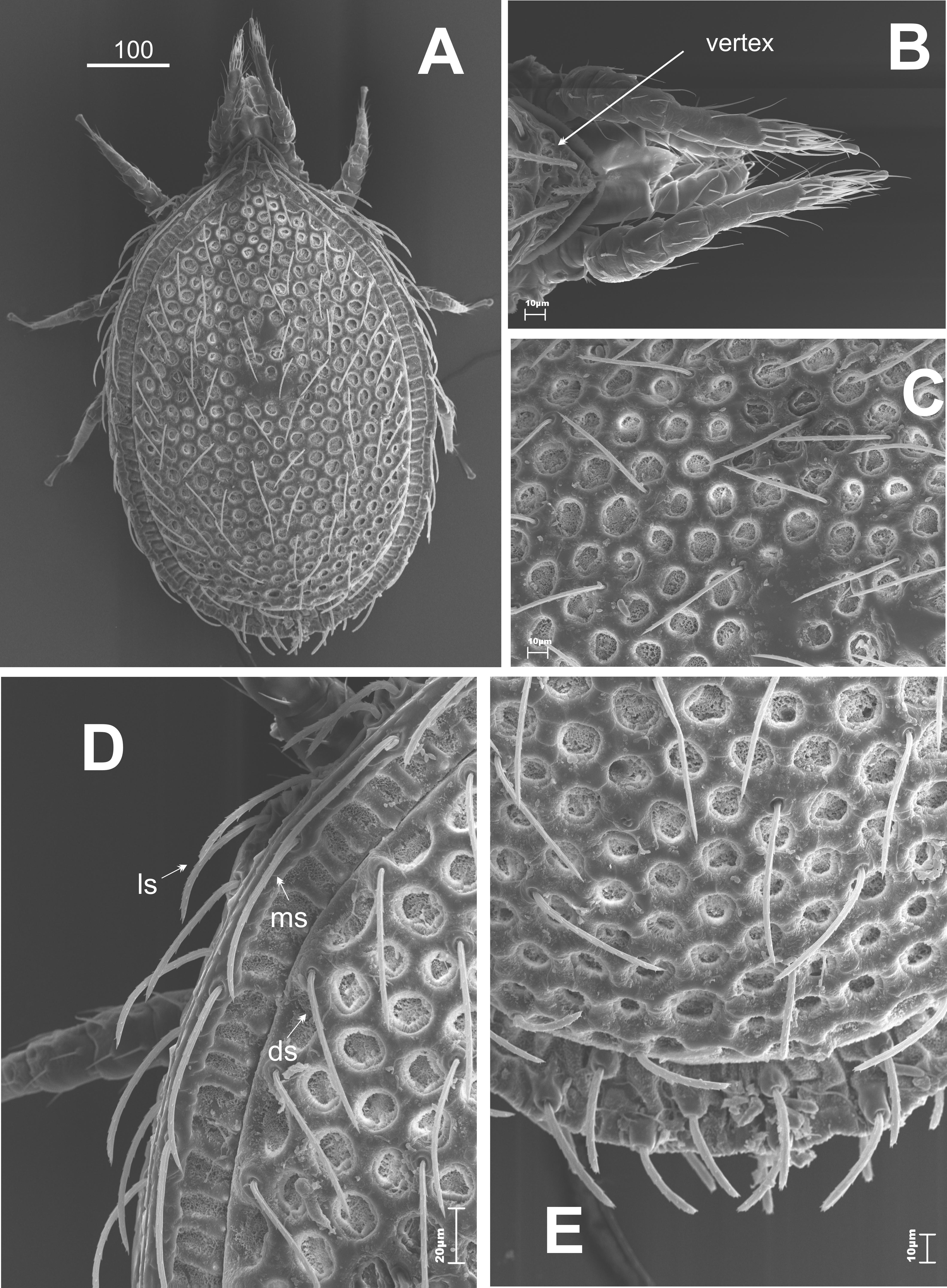


The marginal setae are similar in length (>40 μm) to the dorsal ones, located on a narrow thickening of chitin, which lies along the external edge of the shield (Figures 1D and 2B); at the posterior part of the body these setae are significantly shorter and located on cylindrical elevations.
The dorsal shield is flat, which can be easily seen from the side view (Figure 2C). The vertex is lowered. There are two pairs of long setae on it (Figure 2D). There are tiny thorns / spikes on the marginal shield – the function is unknown. These structures can be easily seen and they are visible at the posterior part of the idiosoma (Figure 3).


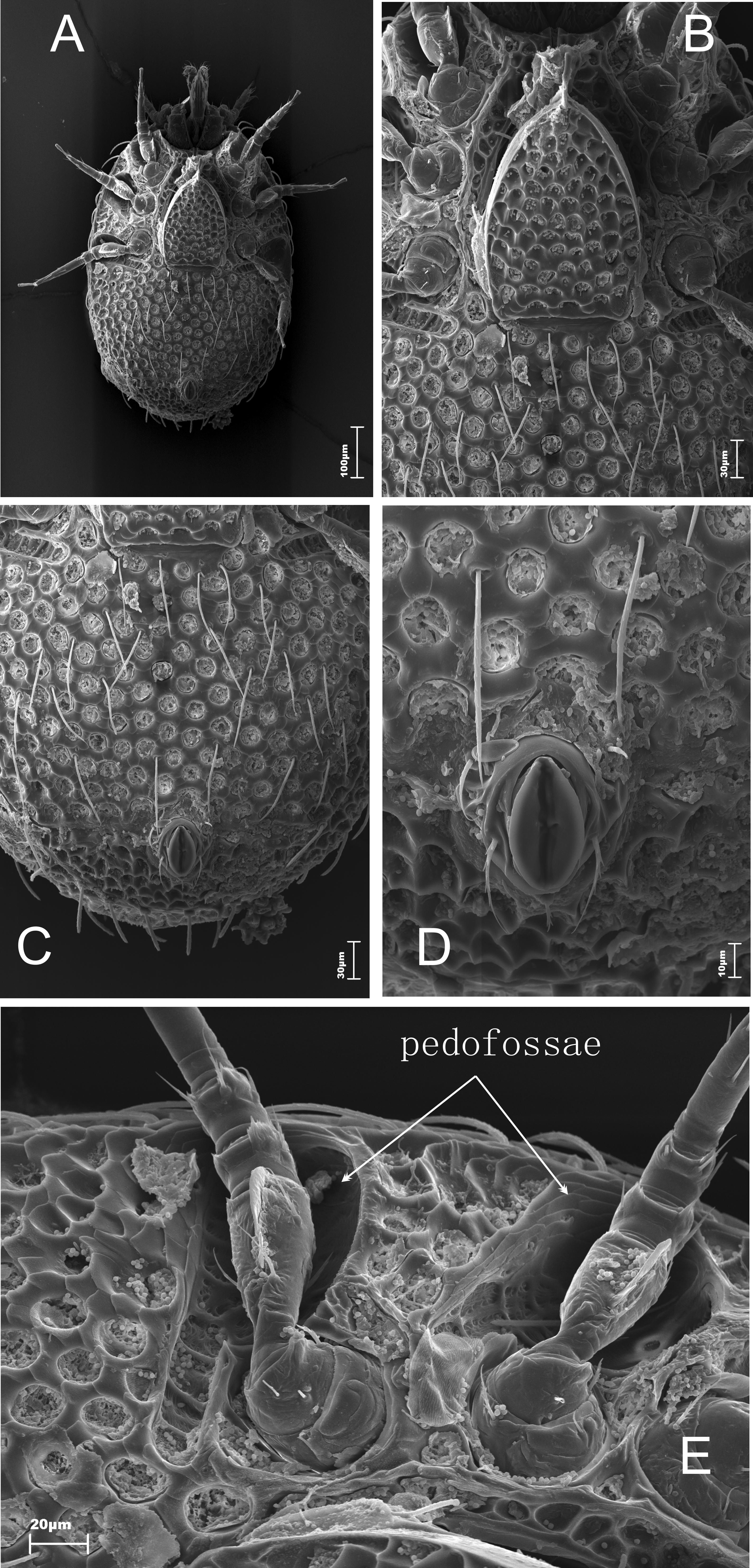
The ventral side (Figures 4A and 4C) is slightly convex, type of sculpture, length and shape similar to the dorsal side (Figure 4A). The large sculptured genital shield with a long appendix at the front occupies the majority of the intercoxal region (Figure 4B). Four pairs of needle-like sternal setae are placed at the sides of the genital shield. They are of various lengths (St1 15 μm, St2 17 μm, St3 28 – 34 μm). The last pair are the longest (St4 52 – 62 μm). In females setae St5 are reduced. The peritreme has a short poststigmal sector. The stigma is situated between the coxae of the second and the third pair of legs. The prestigmal sector is elongated and bent. Cavities for legs are present. The pedofossae for the fourth pair of legs are located transversally and they do not overlap with the opisthosoma (Figures 4A and 4E). The ventral setae (usually 12-13 pairs) are long linear and serrated. They form some transversal rows. The two pairs of those setae are placed a little bit below the back edge of the genital shield (Figure 4C). There are two pairs of needle-like adanal setae. The second pair is significantly longer (Ad1 23 μm, Ad2 31 – 35 μm) and gently serrated (Figure 4D). The single postanal seta (Pa) is very short (10 μm), needle-like, and gently serrated.

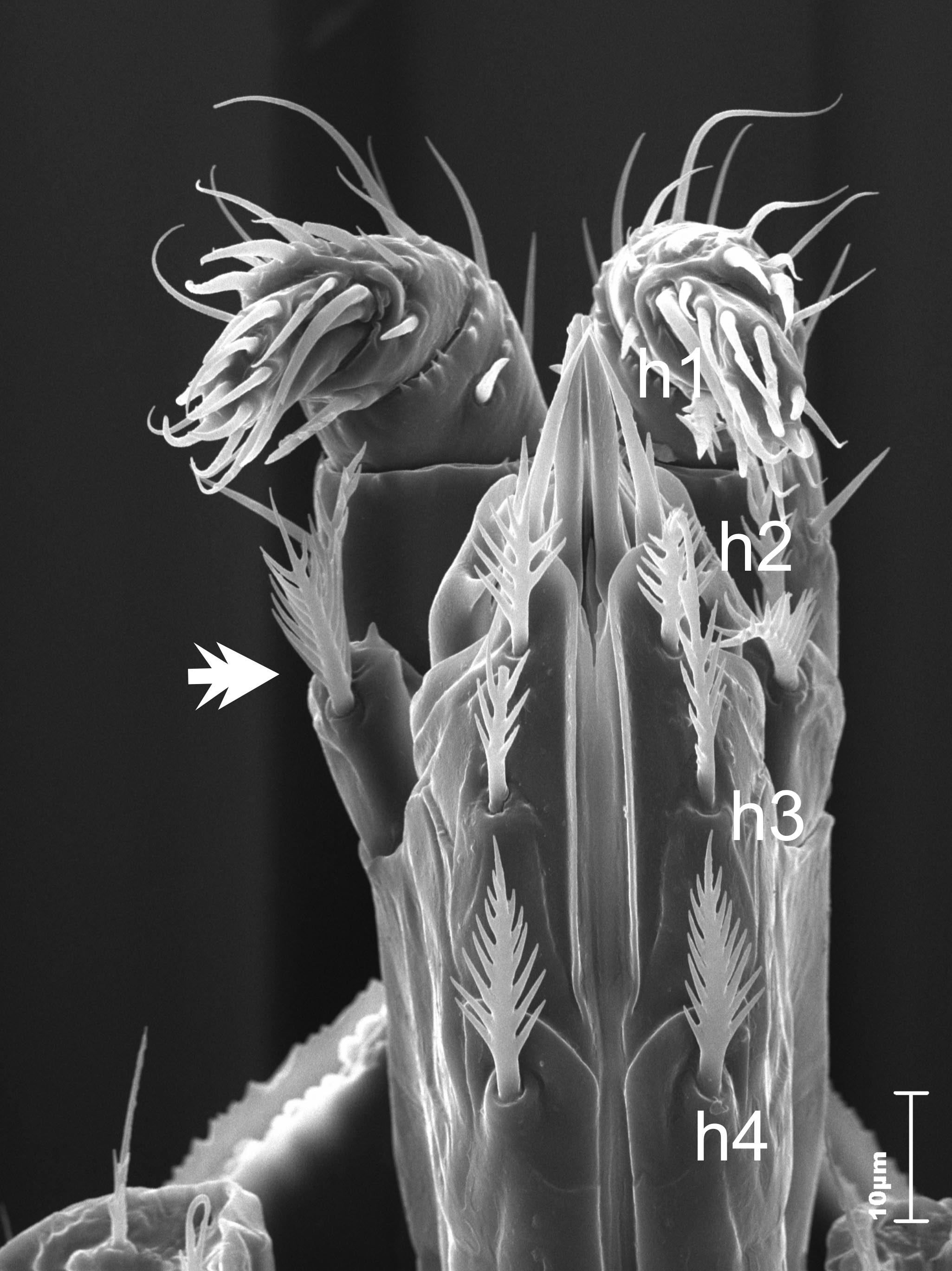
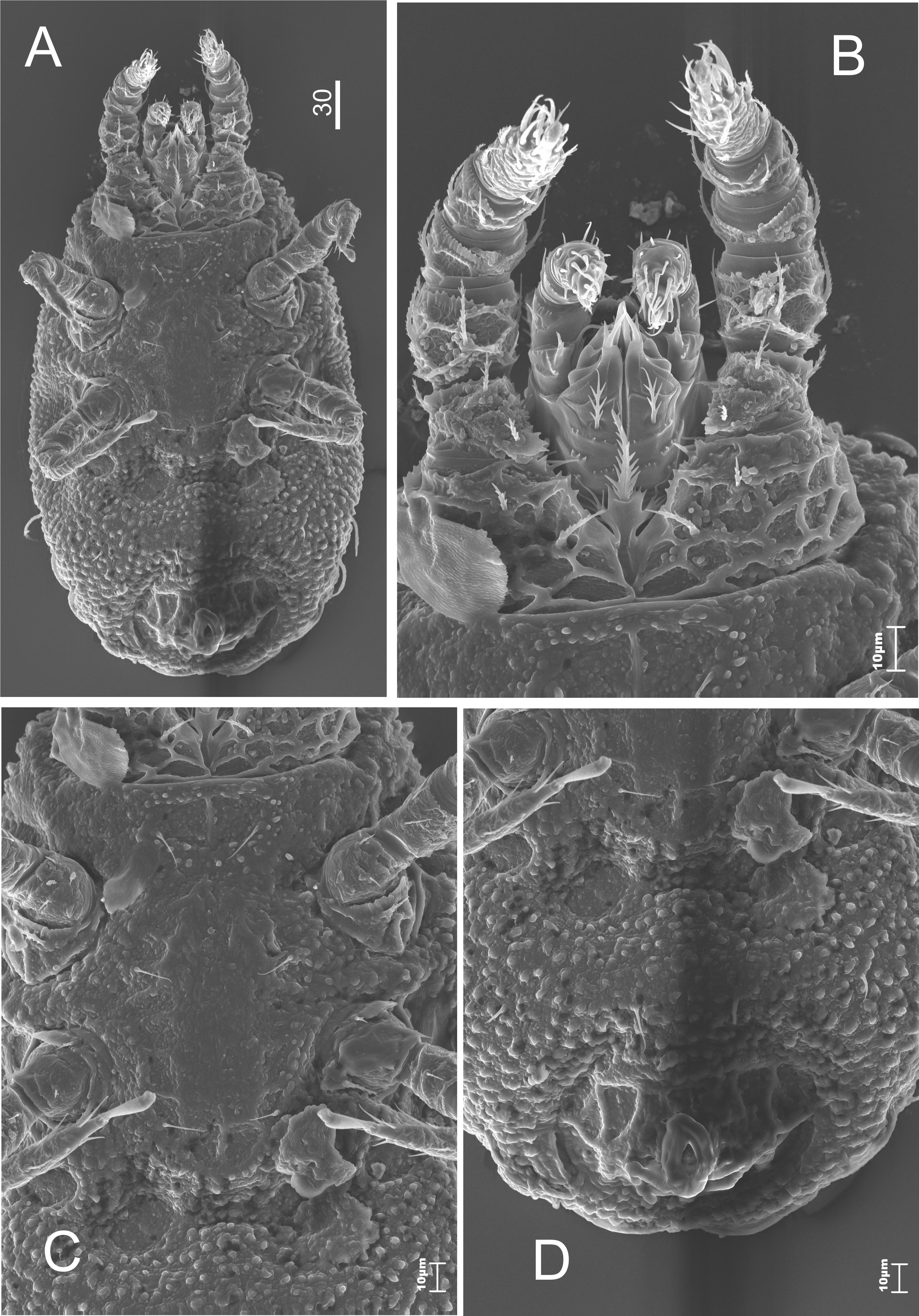
The tritosternum has a narrow base (covered by coxa of the first pair of legs). It also has a strongly feathery, single lacini. The corniculi resembles the rutellum of Opilioacarida. The hypostomal groove is narrow with some teeth (oligodontic type). The hypostomal setae: h1 needle-like (about 20 μm), h2-h4 highly serrated (15, 20, 20 μm, respectively). The trochanters of palps have a pair of feathery setae (Figure 5). The chelicerae are robust.

Male.
The dorsal side (Figure 6). The sculpture and chaetotaxy of the dorsal side are very similar to those in females (compare: Figure 1 and Figure 6).


The ventral side (Figure 7). The sculpture of the ventral side is like the one found in females. The small oval genital shield is located centrally, in the intercoxal region (at the level of the third pair of legs). There are five pairs of needle-like sternal setae (st1-st5) of different length (Figures 7A and 7B): st1-st3 short (8, 15, 26 μm), st4 long (32 – 34 μm), st5 short (26 μm). At the sides of the first three pairs of the sternal setae one can notice characteristically sculptured areas that surround pores. The chaetotaxy of opisthosoma is the same like in females.

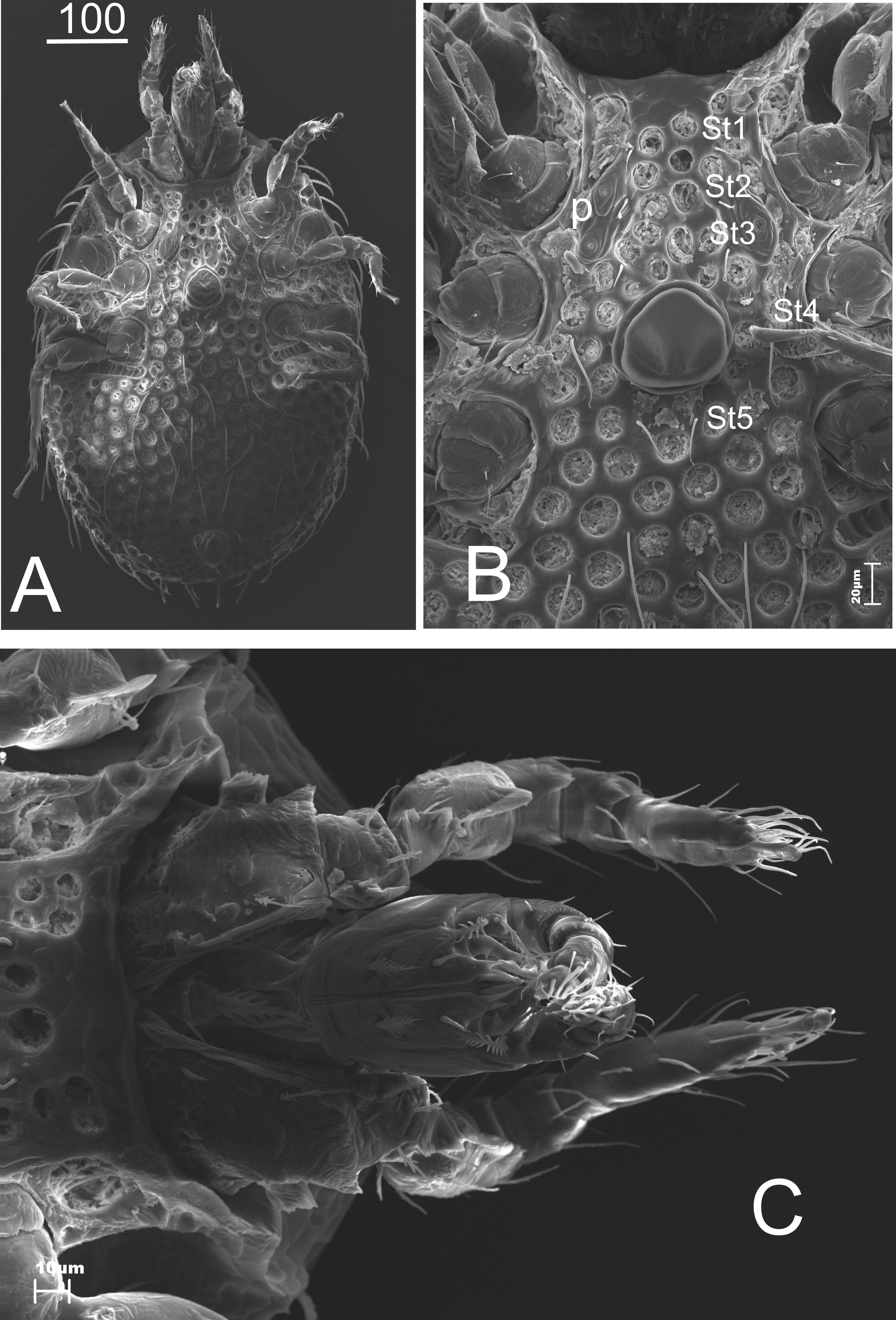
There is a gnathosoma. Clearly noticeable sexual dimorphism. Males have reduced h2 setae and – unlike in females – h3 setae are long needle-like and not serrated (Figure 7C).
Deutonymph.
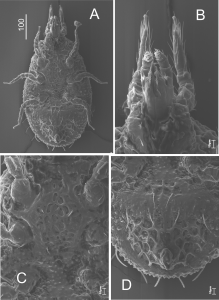
The dorsal side (Figure 8). The body is oval with a small triangular vertex at the front (Figures 8A and 8B). The dorsal shield is big and highly sculptured. It is surrounded by the marginal shield, which has a sclerotized narrow band along the edges. The dorsal setae are long (38 – 43 μm) linear and serrated (Figure 8D).

The dorsal part is only partly sclerotized. The marginal shield (very narrow) and the dorsal shield are separated by soft pleura (Figure 8E).
The ventral side (Figures 9B-9D). The sternal shield is intensively sculptured. It has a shape of an elongated cup. The sculpture is made of oval caveoli of various diameters. There are five pairs of needle-like sternal setae and of similar length (32 – 39 μm). Cavities for legs are present, but they are not as arched, as in adults. The peritreme is relatively long, with a short, straight poststigmal sector (Figure 9A). The ventro-anal shield is big oval and has four pairs of long needle-like setae (48 – 51 μm) in a row, placed closely at the front edge, and there are also two pairs of setae in the middle part. The anus is small and round, with two pairs of needle-like setae. The structure of the anal region does not imply phoresy.All ventral shields of newly molted deutonymphs adhere closely to each other. When a mite grows, they begin to grow apart and the space between them is filled by soft pleura. At the last stage, the deutonymph grows to the size of an adult and develops into a male or female.


Protonymph.
The dorsal side (Figure 10). The idiosoma is oval with a small triangular vertex at the front. The highly sculptured dorsal shields are separated. The sculpture is made of polygons of various size. They are made of thickenings of chitin. There are large podonotal shields in the anterior part of the idiosoma. The five pairs of the dorsal setae are long. They are arranged in two longitudinal rows in the middle of the shield (Figures 10A and 10C). Below there are two triangular mezonotal shields, which are free from the setae. The large oval pygidial shield is not covered by the setae. The other dorsal setae are placed on the chitinous reinforcements at the base, at the soft pleura (Figure 10E). A row of 11 submarginal setae runs at the sides of the podonotal shield, mesonotal shields, and pygidial shield. Two pair of setae are placed over the front edge of the pygidial shield.


The ventral side (Figure 11). The trapezoid-like sculptured sternal shield reaches slightly beneath the coxa of the third pair of legs. There are three pairs of short needle-like setae. The ventro-anal shield has a polygonal sculpture with a large mesh. The other parts of the opisthosoma and the sides of the body, like in larvae, are covered by characteristic granulation. The ventral setae are long needle-like and slightly serrated (Figure 11D). The first pair of them is located at the border of the podosoma and opisthosoma, the two other pairs are located over the front edge of the ventro-anal shield. The anus is oval, slightly elongated, and located at the back area of the shield.

There are two pairs of hypostomal setae: the h1-h3 is smooth, the h4 is feathery. The two pairs of the serrated setae are located on the flattened coxae I, and the first pair of the setae is longer.
Larva.
The dorsal side (Figure 12). The tritosternum has a narrow base, covered by coxae of the first pair of legs, with notched front edge and feathery lacini, which becomes more and more narrow towards the top. There are two pairs of hypostomal setae. The h1 is smooth, the h4 is feathery. One can notice a pair of serrated setae on each of the flattened coxae of the first pair of legs; the first seta is two times longer than the second one.

The ventral side (Figure 13). The sternal shield is weakly sclerotized and smooth. It overlaps with the opisthosoma. There are three pairs (st1-st3) of needle-like sternal setae. The ventro-anal shield is oval with polygonal sculpture and big mesh. The rest of the opisthosoma and the sides of the body are covered with characteristic granulation. The four ventral setae are short and needle-like (Figure 13D). The oval anus is located at the posterior edge of the shield, without circumanal setae.

Notes — The number of sternal setae changes considerably during the ontogenesis period, depending on the developmental stage. Larvae and protonymphs have three pairs of the setae (St1-St3), though larvae the sternal shield is divided into two parts (one on the front part and two pairs on the back part). Deutonymphs have 5 pairs of sternal setae on the long sternal shield (St1-St5). This number is the same in adult males, whereas adult females have no St5 setae.
A similar situation can be observed in the case of the hypostomal setae. Larvae have only two setae (h1 and probably h3). Protonymphs have three pairs of setae on the hypostome (there are no characteristic smooth setae h1). Females have all pairs of the hypostomal setae (h1-h4), whereas in males the setae h2 is reduced, and there are also changes in the shape of the setae h3.
Locus typicus: Thuringia (Thüringen), Germany.
Trematurella elegans was described by Kramer (1882) from Thuringia (central Germany). The species was found in many other countries, not only in Europe. At present, the species occurs in Germany, France, Great Britain, Sweden, Finland, Poland, Czech Republic, Slovakia, Ukraine, Hungary, Greece, Iran (see Taxonomy) (Figure 14). In the majority of these countries the species was found in a few locations. Mašan (2001) found this species mostly in rare locations, mainly in the western and south-western parts of Slovakia.


The authors of this study have tried to obtain all information about the distribution of this species from the available literature, though due to the fact that the species has been found so far in very few countries it is possible that T. elegans has much broader range of occurrence than that presented here. This may also stem from the fact that the species is rare and it occurs in low abundance. Thus, the presence of the species in those countries in which it has not been found yet cannot be excluded. This can also be said about the Balkans and the area of the Mediterranean. The presence of this species in Greece and Iran suggests much broader range of occurrence than that presented in the literature. For this reason the information about the distribution of T. elegans presented in this study should be considered as a summary of the accounts from the literature on the topic, and not as a new description of the current distribution of the species. The strong preference of the species for bark galleries habitats and occurrence in dead wood of different species of trees in different types of forests suggest that T. elegans may have come to the areas of central and northern Europe already in the Boreal or in a warmer Atlantic period from the area of the area of Mediterranean.
The detailed analysis of distribution of T. elegans in Poland (Figure 15) showed that the species is uniformly distributed in the country, except West Pomerania (NW). The range of occurrence is situated on the meltwater channel of Noteć, which is a part of Eberswalde –Torun meltwater channel. Such a distribution suggests that this species did not manage to colonize the area covered by the last glaciation (Wechselian, syn=Würm), which took place in the area of Poland in Pleistocene and finished c.a. 12,000 years ago (Stankowski 1978; Kondracki 2011). After the Mindel (=Elsterian, 730 and 430 years ago) glaciation and the Saalian (=Riss), which took place between 300 and 170 thousands years ago (Stankowski 1978; Kondracki 2011), this species could disperse on the ice-free areas from two directions – from south-west (through the present Germany territory) and from south-east (through the present Slovakia and Ukraine territory). However, one cannot exclude that the species survived the glaciations in this part of Poland, which was not under the glacier (namely, southwards from the line of Cracow – Przemyśl), and it started the expansion from there to the north.

The research into habitat preferences of T. elegans conducted in Poland is based on 7,476 samples, from 35 types of habitats and microhabitats (Table 1), which show more precisely habitat preferences of this species.
The frequency of the species in the samples was low (2.47%). Therefore, the occurrence of the species can be regarded as accidental (F<5%). The percentage of T. elegans in the fauna of Uropodina of Poland is low (0.87%). Hence, the species is regarded as rare and not numerous.
Trematurella elegans can be classified as a typically forestal species. As it is shown in Table 1, it avoids open and non-woody habitats (Table 1). It has been found neither in the upper subalpine of spruce forests, mugo pines, or other scrubs, nor in anthills. The highest frequency has been recorded in natural and planted spruce forests of the Polish part of the North (Middle) European Plain. The species often inhabits larch stands and yew-tree stands. Furthermore, hornbeam forests, mixed deciduous forests, oak-woods and fir-beech forests can be regarded as favorable habitats. In these forests T. elegans can be found under the tree bark (frequency c.a. 25%), in litter, soil, dead wood, and in tree hollows. However, the frequency is many times lower in those habitats than in the most frequent ones. Interestingly, spruce-containing woods are inhabited with lower frequency than other coniferous woods (Table 1).


The appearance of the life stages of T. elegans in various seasons was analyzed on the basis of 7,190 samples (Table 2). The peak of the abundance of T. elegans occurs in June (Figure 16). The number is also high in samples collected in May and August. However, it was very low at the beginning of the phenological season in April and the end of it (December).

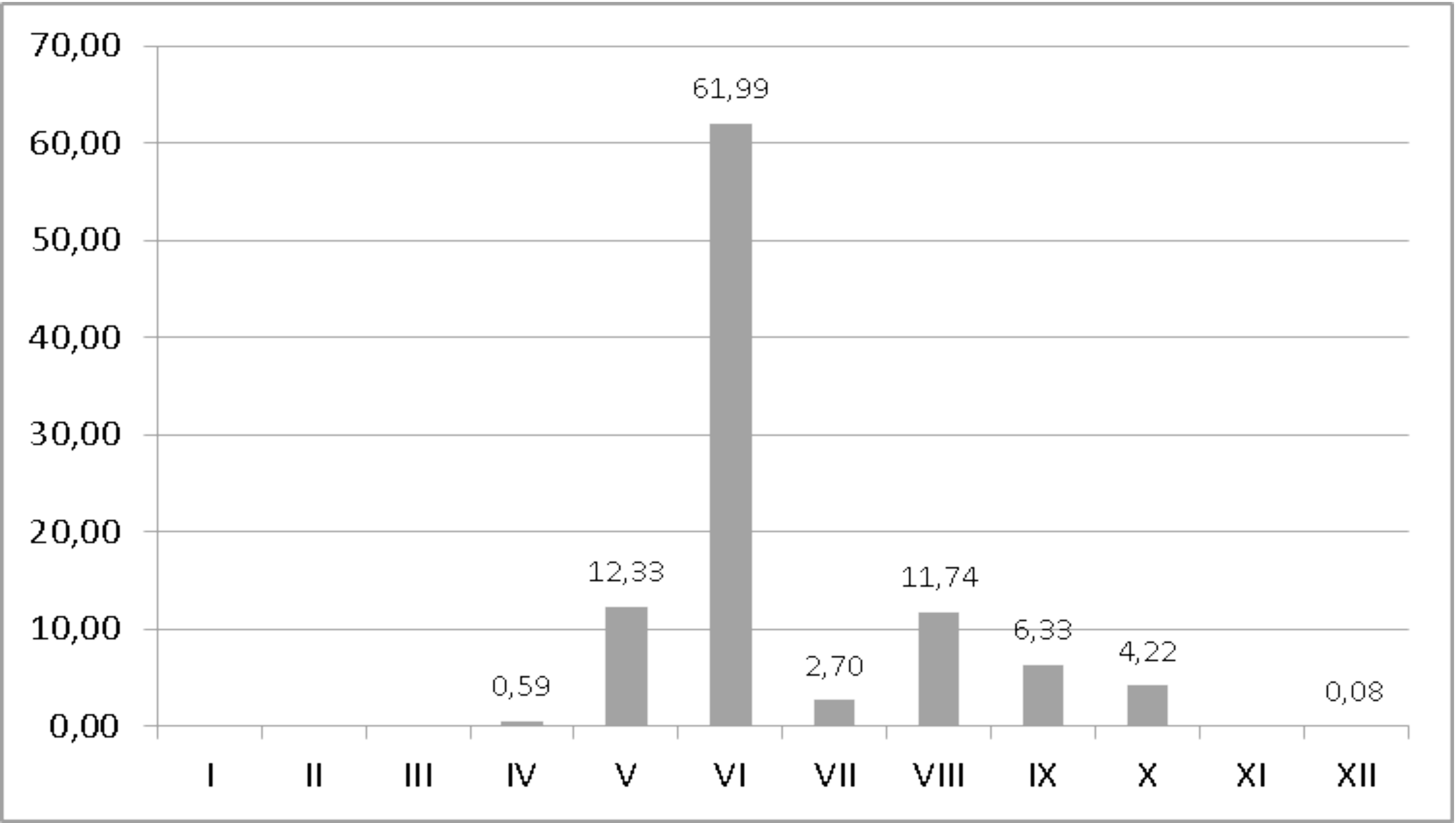
The constant presence of adults and deutonymphs in the collected samples was recorded between April and October (Table 2). Single male specimens were found in December, which suggests that these mites overwinter as adults and deutonymphs hidden under bark of trees. Unfortunately, we do not have enough samples collected in winter to support this hypothesis. The diagram shows (Figure 16) that the presence of larvae and protonymphs is limited in time and has two peaks of abundance, i.e. the first in June and the second in October. The larvae were found in May, September, and October, whereas the protonymphs occurred between May and August, and in October.


The analysis of ventrical distribution of T. elegans is based on 7,506 samples collected at various altitudes in Poland (Table 3). The analysis shows that T. elegans is a typical lowland species, which does not inhabit altitudes over 550 m above sea level. The most optimal altitude for the species rangesbetween 100 and 450 m a.s.l. (Table 3).

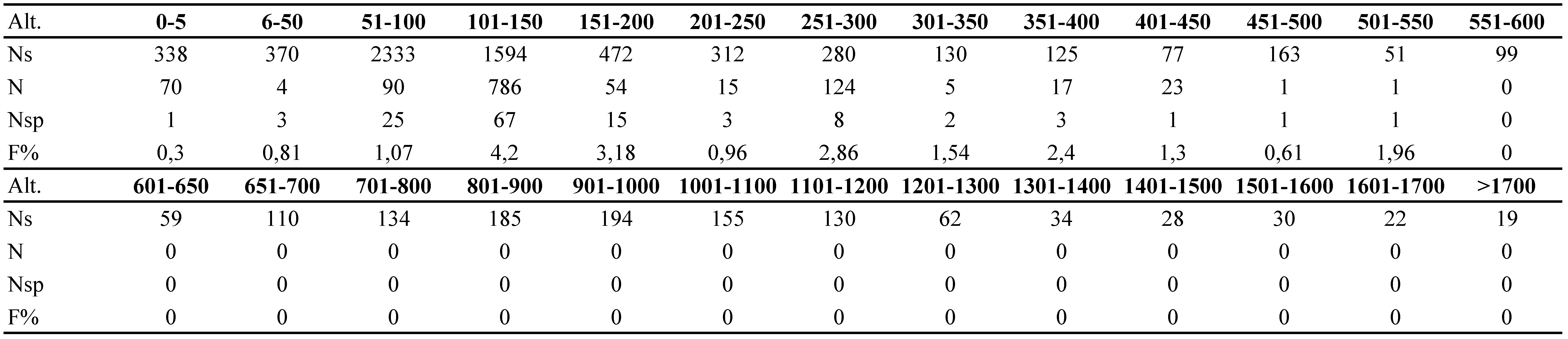
The results shown in this article support the earlier observations of other authors who discuss the distribution and ecology of T. elegans. Karg (1989) described T. elegans as a European species, associated with litter and soils of leafy forests, bark of trees, and with anthills. In the catalogue by Wiśniewski and Hirschmann (1993) T. elegans is described as a species occurring in various environments like mosses, holes in trees, dead wood, forest bed, and anthills of Formicidae (Hymenoptera). However, since the observations mentioned above were rather superficial, it was difficult to establish the preferred habitats of the species on the basis of those studies. These authors described the distribution of the species as northern- and central- European (Wiśniewski and Hirschmann 1993). Mašan (2001) describes the geographical distribution of T. elegans in a similar way. He regards this species as a detricol, which is associated with various types of substrates, such as soil, litter, decaying plants, tree bark, and dead wood. Moreover, he found the species in nests of Turdus philomelos C. L. Brehm, and Clethrionomys glareolus (Schreber), and in anthills. The last habitat mentioned above was also mentioned by Karg (1989). The studies published by Mašan (2001) also show that the distribution of the species in Slovakia is not regular, in the whole area of the country, similarly like in Poland (Błoszyk 1999). Furthermore, the vertical distribution and preferred altitudes are similar in both countries. Mašan did not find T. elegans over 600 meters a.s.l. and he claims that the optimal altitude is lower than 400 meters a.s.l. Fenda and Ciceková (2009) found this species in the south-western Slovakian oak-hornbeam forest in the Little Carpathian Mountains. Kontschán (2008) found T. elegans in Hungary, but he did not include any information about the habitat preferences. The species was also described in a similar way by Kontschán (2013). The authors believe that the species Trichouropoda graeca Kontschán, 2003, which he found in Greece, should be considered as a synonym of T. elegans due to the imprecise description and very schematic drawing, which does not show any significant differences between the two species. The differences probably stem from the geographical variation of T. elegans. Also Kaczmarek et al. (2011) points out that T. elegans is a dominant species in tree hollows in Tilia cordata. Lehtinen (1987), followed by Huhta (2016), found this species in dead wood and anthills in Finland. This species had been also found in anthills by Wiśniewski and Hirschmann (1991), but their findings do not conform to the results presented above (Table 1).
The results of the research presented here confirm that T. elegans is a European species, which inhabits mainly old forests. This probably stems from the type of the tree stands in which the species occurred most frequently (Table 1). This can be also observed in the case of the Białowieża Forest (NE Poland) – the largest remaining parts of the primeval forest in Europe, where the high frequency and abundance of T. elegans was recorded (Błoszyk – unpublished data). One of the most important findings concerning the ecology of the species is the high frequency of T. elegans in samples collected under tree bark (25%). This in turn suggests that T. elegans is a typically sub-bark species. It seems that this species can be found more often under bark than in litter, soil or dead wood, where it has been usually found so far (Lehtinen 1987; Karg 1989; Wiśniewski and Hirschmann 1993; Kaczmarek et al. 2011; Huhta 2016). The results of the analysis of phenology of T. elegans indicate that the species probably winters under tree bark as deutonymphs and adults. However, this hypothesis requires further research.
A compilation of both the data from the existing literature concerning biology, ecology, and distribution of T. elegans and the new more detailed data presented here is not only a succinct summary of the information about this species, but it also allows to analyze the history of dispersion of the species in Europe, especially in Poland, after the glaciations. The glaciations affected the range of many species of soil fauna, including mites. However, most of the studies focus on insects (e.g. Lunt et al. 1998; Fattorini and Ulrich 2012) and springtails (e.g. Werner and Fiera 2009). Little is known about the influence of the glaciation on distribution and range of occurrence of soil mites, though some attempts to solve this problem have been already made (Błoszyk et al. 2017). This of course stems from the lack of fossils. Thus, the reconstruction of the migration routes of mites after the glaciation can be possible only on the basis of the knowledge about the development of the environment that is inhabited by particular species, and on the basis of the knowledge of ecological needs of the species. Among Uropodina there are many species which have their northern range of occurrence in Poland (Błoszyk 1999), and T. elegans is also among them. The analysis of the habitat preferences of the species suggests that it reached the northern Europe relatively recently, about few thousand years ago. It probably happened in the boreal interval (c.a. eight thousand years B.P.) because at that time forests were formed in this area, which could provide environmental conditions favorable for T. elegans. The presence of the species in southern Europe (Greece) and in Iran is hard to explain. Perhaps these two countries are old relic areas. There is no doubt that this species is not typical for the Mediterranean. It is possible that like in the case of Trachytes aegrota (C.L. Koch, 1841), T. elegans has its center of occurrence in central Europe, and it migrated from this region to the North and South (Błoszyk 1999; Athias-Binche 1981a,b; 1982a,b).
The microscope scans were made in Electron and Confocal Microscope Laboratory. The authors are also deeply indebted to Prof. Jerzy Gutowski for his consent to collect samples from soil, dead wood, and from bark galleries found in the national reserve Białowieski National Park. We are also grateful to Ms Joanna Gogol for her help in preparing the manuscript.
Athias-Binche F. 1981a. Différent types de structures des peuplements d'Uropodides édaphiques de trois écosystèmes forestiers (Arachnides: Anactinotriches). Acta Oecol.- Oecol. Gen., 2: 153-169.
Athias-Binche F. 1981b. Écologie des Uropodides édaphiques (Arachnides: Parasitiformes) de trois écosystèmes forestiers. 1. Introduction, matérial, biology. Vie Milieu, 31: 137-147.
Athias-Binche F. 1982a. Écologie des Uropodides édaphiques (Arachnides, Parasitoformes) des trois écosystèmes forestiers. 3. Abondance et biomasse des microarthropodes du sol, facteurs du milieu, abondance et distribution spatiale des Uropodides. Vie Milieu, 32: 47-60.
Athias-Binche F. 1982b. Écologie des Uropodides édaphiques (Arachnides: Parasitiformes) des trois écosystèmes forestiers. 4. Abondance, biomasse, distribution verticale, sténo-et eurytopie. Vie Milieu, 32: 159-170.
Berlese A. 1888. Acari, Myriapoda, et Scorpiones hucusque in Italia reperta. Padova, 49. 15 text pages + Plates 1-10.
Berlese A. 1913. Acaroteca Italica. Fasc. 1 and 2. In: Tipografia M. Ricci, Firenze (Eds). p. 221.
Berlese A. 1917. Centuria seconda di Acari nuovi. Redia, 12(I e II): 144-145.
Błoszyk J. 1983. Uropodina Polski (Acari: Mesostigmata) [PhD Thesis]. Poznań: Adam Mickiewicz University. pp. 543.
Błoszyk J. 1984. Pionowe zróżnicowanie fauny Uropodina (Acari) Polski. Prz. Zool., 28: 69-71.
Błoszyk J. 1999. Geograficzne i ekologiczne zróżnicowanie zgrupowań roztoczy z kohorty Uropodina (Acari: Mesostigmata) w Polsce. I. Uropodina lasów grądowych (Carpinion betuli). Poznań: Wyd. Kontekst. pp. 245.
Błoszyk J., Adamski Z., Napierała A., Dylewska M. 2004. Parthenogenesis as a life strategy among mites of the suborder Uropodina (Acari: Mesostigmata). Can. J. Zool. 82: 1503-1511. doi:10.1139/z04-133 ![]()
Błoszyk J., Athias-Binche F. 1998. Survey of European mites of cohort Uropodina. I. Geographical distribution, biology and ecology of Polyaspinus cylindricus Berlese, 1916. Biol. Bull. Poznan. Zool., 32: 89-102.
Błoszyk J., Krysiak D. 2000. Uropodina (Acari: Mesostigmata) rezerwatu "Cisy Staropolskie im. Leona Wyczółkowskiego" w Wierzchlesie. Parki Nar. Rez. Przyr., 19(2): 115-121.
Błoszyk J., Krysiak D., Napierała A., Bajaczyk R. 2002a. Materiały do znajomości Uropodina (Acari: Mesostigmata) wybranych rezerwatów położonych w obszarze parków krajobrazowych województwa wielkopolskiego. Biul. Park. Krajobraz. Wielkop., 8: 61-72.
Błoszyk J., Krysiak D., Napierała A., Dylewska M. 2006. Can soil fauna undergo synanthropisation? In: Gabryś G., Ignatowicz S. (Eds). Advances in Polish Acarology. Warszawa: SGGW. p. 26-37.
Błoszyk J., Krysiak D., Napierała A., Markowicz M. 2002b. Ocena stanu środowiska glebowego wybranych rezerwatów leśnych województwa wielkopolskiego w oparciu o zgrupowania roztoczy z podrzędu Uropodina (Acari: Mesostigmata). In: Pułyk M., Tybiszewska E. (Eds). Raport o stanie środowiska w Wielkopolsce w roku 2001. Poznań: WIOŚ w Poznaniu. p. 21-31.
Błoszyk J., Książkiewicz-Parulska Z., Adamski Z., Napierała A. 2017. Influence of Pleistocene glaciation on the distribution of three species of Labidostomma in Europe (Acari: Labidostommatidae). Syst. Appl. Acarol., 22(6): 841-857. doi:10.11158/saa.22.6.9 ![]()
Błoszyk J., Markowicz M., Labijak B., Skwierczyński F., Napierała A. 2015. Microgeographic diversity of Uropodina (Acari: Mesostigmata) communities in dead wood and tree hollows. Redia, XCVIII: 3-12.
Błoszyk J., Napierała A. 2004. Roztocze z podrzędu Uropodina (Acari: Mesostigmata) Wyżyny Krakowsko-Częstochowskiej. In: Partyka J. (Eds). Zróżnicowanie i przemiany środowiska przyrodniczo-kulturowego Wyżyny Krakowsko-Częstochowskiej. T. 1. Przyroda. Ojców: Ojcowski Park Narodowy. p. 285-290.
Błoszyk J., Nowak M., Mokrowiecka A. 2013. Rozmieszczenie roztoczy z podrzędu Uropodina (Acari: Mesostigmata) w Gorczańskim PN. In: Kalinowska-Szymczak, A. (Eds). Kalejdoskop GIS. Tom 2. Warszawa: Esri Polska. p. 88-89.
Fattorini S., Ulrich W. 2012. Spatial distributions of European Tenebrionidae point to multiple postglacial colonization trajectories. Biol. J. Linn. Soc., 105: 318-329. doi:10.1111/j.1095-8312.2011.01797.x ![]()
Fenda P., Ciceková J. 2009. Soil mites (Acari: Mesostigmata) of oak-hornbeam forest in NR Katarínka, Southwest Slovakia. In: Tajovský K. Schlaghamerský J., Pižl V. (Eds). Contributions to Soil Zoology in Central Europe III. České Budějovice: Institute of Soil Biology Academy of Sciences of the Czech Republic. p. 29-32.
Halbert J.-N. 1915. Clare Island survey. Acarinida. Terrestrial and marine Acarina. Proc. Roy. Ir. Acad. 31B: 45-136.
Halbert J.-N. 1923. Notes on Acari, with descriptions of new species. J. Linn. Soc. Zool., 35: 363-392 + Plates 20-22.
Haller G. 1881. Acarinologisches. Archiv für Naturgeschichte, 47: 182-193 + Plate IX.
Hermann J. 1804. Mémorie Aptérologique. Strasbourg : Levraut. 136 pp. + Plates I-IX.
Hirschmann W., Zirngiebl-Nicol I. 1961. Gangsystematic der Parasitiformes. Teil. 4. Acarologie, Schriftenreihe für vergleichende Milbenkunde, 4: 1-41.
Huhta V. 2016. Catalogue of the Mesostigmata mites in Finland. Memor. Soc. Fauna Flora Fenn., 92: 129-148.
Kaczmarek S., Marquardt T., Faleńczyk-Koziróg K. 2011. Diversity of the Mesostigmata (Acari) in tree-hollows of selected deciduous tree species. Biological Lett. 48: 29-37.
Karg W. 1989. Acari (Acarina) Milben, Unterordnung Parasitiformes (Anactinochaeta). Uropodina Kramer, Schildkrötenmilben. Tierwelt Deutschlands 67: 1-203.
Kazemi S., Kontschán J. 2014. Three new records of the superfamily Uropodidea (Acari: Mesostigmata: Uropodina) from Iran, and a key to the known Iranian species of Uropodidea Persian J. Acarol., 3(1): 9-16.
Koch C.-L. 1835. Deutschlands Crustaceen, Myriapoden und Arachniden. Ein Beitrag zur Deutschen Fauna. Vol. 2. Regensberg: Herrich-Schäffer. 42 un-numbered text pages + 24 plates.
Koch C.-L. 1839. Deutschlands Crustaceen, Myriapoden und Arachniden. Ein Beitrag zur Deutschen Fauna. Vol. 27. Regensberg: Herrich-Schäffer. 25 un-numbered text pages + 24 plates.
Koch C.-L. 1841. Deutschlands Crustaceen, Myriapoden und Arachniden. Ein Beitrag zur Deutschen Fauna. Vol. 32. Regensberg: Herrich-Schäffer. 32 un-numbered text pages + 24 plates.
Koch C.-L. 1847. System der Myriapoden, mit den Verzeichnissen und Berichtigungen zu Deutschlands Crustaceen, Myriapoden und Arachniden Heft 1–40. Regensburg: Putset. 270 pp. + plates I–X.
Kondracki J. 2011. Geografia regionalna Polski. Warszawa: Wydawnictwo Naukowe PWN. pp. 441
Kontschán J. 2002. Adatok Komárom-Esztergom megye korongatka (Acari: Uropodina) faunájához. Komárom – Esztergom Megyei Múzeumok Közleményei, 9: 345-352.
Kontschán J. 2003. Data of the Uropodina (Acari: Mesostigmata) of Greece and Malta. Annls. hist.-natur. Mus. natn. hung., 95: 185-191.
Kontschán J. 2008. Magyarország korongatkái (Acari: Mesostigmata: Uropodina). Állat. Közlem., 93(1): 3-15.
Kontschán J. 2013. Uropodina mites of the Balkan Peninsula (Acari: Mesostigmata). Opusc. Zool. Budapest, 44(suppl. 1): 97-131.
Kramer P. 1882. Ueber Gamasiden. Arch. Naturgesch., 48: 406-407.
Lehtinen P.-T. 1987. Association of uropodid, prodinychid, polyaspidid, antennophorid, sejid, microgynid, and zerconid mites with ants. Entomol. Tidskr., 108: 13-20.
Lunt D., Ibrahim L.K., Hewitt G.M. 1998. mtDNA phylogeography and postglacial patterns of subdivision in the meadow grasshopper Chorthippus parallelus. Heredity, 80: 633-641. doi:10.1046/j.1365-2540.1998.00311.x ![]()
Mašan P. 2001. Mites of the cohort Uropodina (Acarina, Mesostigmata) in Slovakia. First ed. Annot. Zool. Bot., Bratislava. 320 pp.
Michael A.-D. 1894. Notes on Uropodina. J. R. Microsc. Soc., p. 289-319 + Plates VI, VII.
Müller O.-F. 1776. Zoologiae Danicae Prodromus, seu Animalium Daniae et Norvegicae Indigenarum. Characteres, Nomina, et Synonyma Imprimis Popularum. Copenhagen: Hafniae. 282 pp.
Napierała A. 2008. Struktura zgrupowań i rozkład przestrzenny Uropodina (Acari: Mesostigmata) w wybranych kompleksach leśnych Wielkopolski [PhD Thesis]. Poznań: Adam Mickiewicz University. pp. 188.
Napierała A., Błoszyk J. 2013. Unstable microhabitats (merocenoses) as specific habitats of Uropodina mites (Acari: Mesostigmata). Exp. Appl. Acarol. 60: 163-180. doi:10.1007/s10493-013-9659-9 ![]()
Napierała A., Błoszyk J., Bruin J. 2009. Communities of uropodine mites (Acari: Mesostigmata) in selected oak-hornbeam forests of the Wielkopolska region (Poland). Exp. Appl. Acarol., 49: 291-303. doi:10.1007/s10493-009-9262-2 ![]()
Napierała A., Mrozek K., Błoszyk J. 2006. Akarofauna Ojcowskiego Parku Narodowego na tle pozostałych parków narodowych w Polsce. Prądnik. Prace i Materiały Muzeum im. Prof. W. Szafera, 16: 153-164.
Stankowski W. 1978. Rozwój środowiska fizyczno-geograficznego Polski. Warszawa: Państwowe Wydawnictwo Naukowe. pp. 193.
Trägårdh I. 1942. Über die Trematurini Berlese (Acarina). Zool. Anzeiger, 139: 109-112.
Trӓgårdh I. 1945. Description of Trematurella Tragardh, a new genus of Uropodina (Acarina). Arkiv. f. Zool., 36A(14): 1-10.
Werner U., Fiera C. 2009. Environmental correlates of species richness of European springtails (Hexapoda: Collembola). Acta Oecol., 35: 45-52. doi:10.1016/j.actao.2008.07.007 ![]()
Wiśniewski J. 1979. Gangsystematik der Parasitiformes. Teil. 312. Acarologie: Schriftenreihe für vergleichende Milbenkunde, 25: 37-38.
Wiśniewski J., Hirschmann W. 1991. Ergӓnzungsangaben über Verbreitung und Lebensrӓume der Trichouropoda-Arten (Acarina, Uropodina). Fragm. Faunist., 35(11): 173-178. doi:10.3161/00159301FF1991.35.11.173 ![]()
Wiśniewski J., Hirschmann W. 1993. Katalog der Ganggattungen Untergattungen, Gruppen and Arten der Uropodiden der Erde. Acarologie. 40, 1-220.



2017-12-06
Date accepted:
2018-04-30
Date published:
2018-07-25
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Błoszyk, Jerzy; Adamski, Zbigniew and Napierała, Agnieszka
 Download article Download low definition
Download article Download low definitionDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)