



A putative fossil sejid mite (Parasitiformes: Mesostigmata) in Baltic amber re-identified as an anystine (Acariformes: Prostigmata)
Dunlop, Jason A.1 ; Walter, David E.2 and Kontschán, Jenő3
1✉ Museum für Naturkunde, Leibniz Institute for Evolution and Biodiversity Science, Invalidenstrasse 43, D-10115 Berlin, Germany.
2Department of Biological Sciences, University of of Alberta, Edmonton, Alberta T6G 2E9, Canada; Queensland Museum 4101 and University of the Sunshine Coast, Queensland, Australia, 4556.
3Plant Protection Institute, Centre for Agricultural Research, Hungarian Academy of Sciences, H-1525 Budapest, P.O. Box 102, Hungary.
2018 - Volume: 58 Issue: 3 pages: 665-672
https://doi.org/10.24349/acarologia/20184263ZooBank LSID: 77E87B16-2E25-45C4-942B-A9211E7C4BCB
Keywords
Abstract
Although there are around 11,400 living species of mesostigmatian mites (Acari: Parasitiformes: Mesostigmata) (Beaulieu et al. 2011), this impressive diversity is not reflected in their fossil record. As noted by Dunlop et al. (2014), a possible reason for this is that few mesostigmatan lineages currently inhabit tree bark which, if current ecology reflects ancient ecology, reduced the chance of them becoming preserved as inclusions in amber. Only four fossil species have been formally named: Sejus bdelloides Koch and Berendt, 1854 from Eocene Baltic amber, Dendrolaelaps fossilis Hirschmann, 1971 from Miocene Chiapas (or Mexican) amber, Palaeozercon cavernicolus Błaszak et al., 1995 from a subfossil cave deposit in the USA, and Alecrogamasus stenocornis Witaliński, 2000, again from Baltic amber. In addition to this, there are further examples of fossil mesostigmatans in the literature (see citations in Table 1). Several of them were discovered in association with other arthropods (Lyubarsky and Perkovsky 2012; Dunlop et al. 2013, 2014), but could not be identified beyond the family or genus.

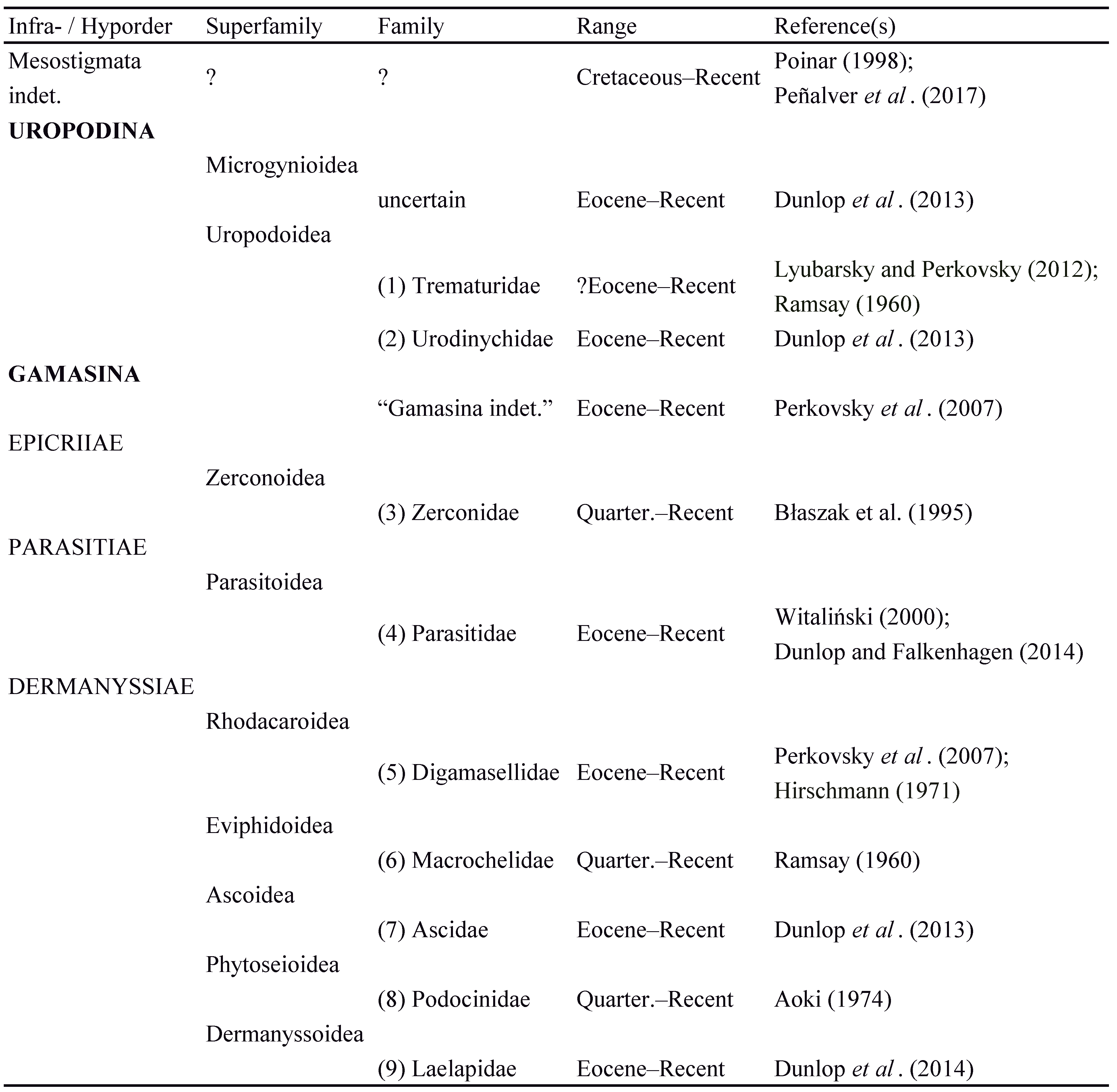
Fossil arachnids are proving to be of increasing importance in calibrating molecular phylogenies by, for example, constraining minimum ages for cladogenesis and offering divergence times between the appearance of stem-group members of a given lineage and crown-group taxa typically assignable to modern genera. Recent examples include Dabert et al. (2010) for acariform mites, Giribet et al. (2009) and Hedin et al. (2012) for harvestmen and Wood et al. (2013) for spiders. Yet before fossils can be used in this way it is important to check the reliability of published attributions. In this context, three principal clades of Mesostigmata are currently recognised: Sejida, Monogynaspida and Trigynaspida. Most of the published mesostigmatan fossils are monogynaspids (Table 1), while Trigynaspida currently lacks a fossil record.
The third group, Sejida is uniquely represented by the historically oldest species: Sejus bdelloides. However, there are several problems associated with this record. In the mid-19th century when this fossil was described the concept of Sejus – sometimes spelled Seius; see synonymy list – was rather loose, the genus name being used for a variety of mesostigmatans. Also, the original line drawings in Koch and Berendt’s (1854) monograph consist of somewhat idealised reconstructions (Figure 1), and are not always reliable guides to the appearance of the actual specimens in the matrix. Thus to clarify whether sejid mites were present in the fossil record, we re-examined the holotype of S. bdelloides (Figures 1, 2) and suggest here that it is neither a Sejida, nor even a Mesostigmata.
The holotype of Sejus bdelloides originates from Baltic amber and lies within in a fairly large piece of dark orange (i.e. partially oxidised) amber, with dimensions of ca. 35 x 15 mm. Two fragments have split off from the main amber piece, but do not contain any inclusions. The specimen is part of the scientifically, and historically, valuable Berendt collection held in the palaeontology section of the Museum für Naturkunde Berlin (MfN). The holotype has the modern repository number MB.A.1634 (for Museum Berlin, Arthropoda). The original label also bears the older collection numbers “117” and “7279”. The amber piece is quite complicated in that it contains numerous syninclusions – most of which are also type and figured specimens – including a cheyletid mite (Acariformes: Prostigmata: Cheyletidae), a clubionid spider and two archaeognath hexapods. Baltic amber is thought to have been deposited in a warm forest environment and is conventionally dated at Eocene (Lutetian), or about 44 – 49 Ma. For a recent overview of this amber deposit and its geological setting see Weitschat and Wichard (2010).
Recognising S. bdelloides from within this diverse assemblage was thus challenging, but we believe it to be the mite towards the middle of the amber piece, tucked in close to the antennae of one of the archaeognaths (Figure 1A). In support of this assertion is the fact that the mite in question bears a small pair of posterior setae, whereby similar setae are indicated both in the original text description “am Hinterrande zwei kurze Borsten”. Furthermore, the notes (“Anmerkungen”) to the original description in Koch and Berendt (1854) state: “Das einzige Exemplar liegt tief in einem etwas dunkelfarbigen Steine und ist überdies noch an der Spitze des Kopfes mit einer undurchsichtigen Blase bedeckt, in welcher auch Rüssel und Taster versteckt sind. Dieses Thierchen gehört bestimmt in die Familie der Gamasiden und den Beinen nach zu gegenwärtiger Gattung.” We translate this as: “The only specimen lies deep in a rather dark stone and is covered from the tip of the head with an opaque bubble which conceals the snout and palps. This little animal certainly belongs in the gamasid family and based on the legs to a living genus.” The specimen figured here has a smooth prodorsal shield, which could appear like a bubble (Figures 1B), and the gnathosoma cannot be seen, thus it fits quite well with Koch and Berendt’s comments. Koch and Berendt (1854) also noted that the first pair of legs was the thickest, which also corresponds well (Figure 1B, 2B) with the specimen we presume to be their holotype.


The specimen was studied and drawn under a Leica stereomicroscope and a Zeiss Axioscope compound microscope; both with a camera lucida attachment. Since the specimen is deeply buried in the now darkened amber, several details in the drawing were visualised with the camera lucida on the compound microscope allowing minute variations in the depth of field to yield more details (e.g. setae); however photographing all of these details with the equipment available was not possible. The specimen was photographed (Figure 1) under a stereomicroscope via the Leica Application Suite software which enables stacks of images to be combined into a final picture. Since the specimen is part of a series of types all within one – now rather fragile – amber piece, it was not permitted to carry out tomographic scans which might have revealed further anatomical details.
Prostigmata Kramer, 1877
Cohort (=Infraorder) Anystina sensu Krantz & Walter, 2009
?Anystoidea Oudemans, 1936, incertae sedis
(Figures 1, 2)
Sejus bdelloides, Koch and Berendt, 1854: 110, pl. 15, fig. 115; Nec Sejus bdelloides [as Seius] Bronn, 1849; Giebel, 1856: 484; Scudder, 1891: 288; nomen nudum according to Oudemans, 1937: 376) Vitzthum, 1943: 730 [as Seius]; Petrunkevitch, 1955: 97 [as Seius]; Selden, 1993: 310 [as Seius]; Walter and Proctor, 1999: 16 [as Seius]; Dunlop et al., 2010: 67; Dunlop et al., 2013: 338.
Material examined ― MB.A.1634 (ex Berendt collection no. 7279). Baltic amber, precise locality not recorded. Palaeogene: Eocene: Lutetian.


Description ― Total body length 656 µm; maximum width ca. 460 µm. Dorsum: Prodorsal shield oval, 364 μm long and 241 μm wide, surface smooth. Prodorsal shield with four pairs of smooth and needle-like setae (potentially representing setae vi, ve, sci and sce), in which case ve and sce are longer (ca. 100 μm ) than ve and sce (ca. 50 μm). Two putative pairs of ocelli situated on membranous cuticle, lateral to prodorsal shield and at level of sce. Four pairs of setae (potentially representing c1, d1, e1 and f1) presented posterior to prodorsal shield. Setae c1 and d1 short (ca. 30 μm) and needle-like, e1 three times longer (ca. 90 μm) and needle-like. Setae f1 absent, only their insertion visible. Setae c2 longer (ca. 130 μm) than other setae on membranous cuticle and situated anterior to c1. Setae h2 or ps1 and not visible. Legs with smooth and simple setae, but all setae and solenida not visible. Claws visible only on tip of leg II, large and with median fleshy pad (empodium).
Remarks ― The original description of Sejus bdelloides does not provide sufficient characters to unequivocally place the fossil in Sejida as it is understood today, simply stating (as noted above) that it’s a gamasid and that, based on the legs, it belongs in the modern genus. Interestingly, Sejus Koch, 1836 was created a few years earlier by one of the describers of the amber inclusion, whereby Carl Ludwig Koch may have been influenced by the apparent posterior setae in the amber specimen (Figure 2B). These are vaguely reminiscent of the condition seen in his (living) type species Sejus togatus Koch, 1836 (see e.g. figures in Hirschmann et al. 1991), although several other mites can show paired posterior setae. However, unlike the dorsally rather setose and pustulate S. togatus, the amber fossil is uniformly plicate except for a prodorsal shield and reveals little in the way of structures consistent with – or diagnostic for – any of the known families of Sejida. A comparative image of Sejus americanus (Banks, 1902) is also included here for reference (Figure 2C) and again the pattern of sclerites and setae here differ markedly from the habitus of the amber inclusion. Sejus bdelloides must therefore be excluded from both Sejida, which now lacks a fossil record entirely, and we see no characters justifying its referral to Mesostigmata in general. The median empodium on leg II also rules out Mesostigmata. We suspect that this amber mite in fact belongs to the acariform suborder Prostigmata. A prodorsal shield with four pairs of setae is visible (Figures 1B, 2B), flanked by what appear to be two pairs of ocelli. This character would also rule out mesostigmatan affinities, as all of these mites should be blind. Note that Koch and Berendt (1854) did not record the eyes, stating “Die Augen sind nicht sichtbar". We are not sure why this discrepancy exists, but the putative eyes are quite hard to see even with modern microscopes (Figure 1B). The legs are relatively stout and tarsus II on the right ends in a pair of claws and empodium (Figures 1B, 2B). Unfortunately the gnathosoma is hidden and the form of the chelicera and palps cannot be determined, but the idiosomal setation is not consistent with any Parasitiformes. That said, Sejus bdelloides does resemble extant members of the cohort Anystina (Walter et al. 2009) and could even be a member of the Anystoidea. Extant Anystoidea are often arboreal, have a similarly shaped body, an anterior dorsal plate with 4 pairs of setae and flanked by two pairs of ocelli (Figure 2B), and similar idiosomal setation. The obscuration of the gnathosoma precludes a definitive identification, but extinct members of Anystoidea have been previously recorded in both Late Cretaceous amber (see Walter and Proctor 2013, Figure 2.12a), Cretaceous and Paleogene Russian amber (Zacharda and Krivolutsky 1985) and Baltic amber by Koch and Berendt (1854) and Sayre et al. (1992). What we interpret as a pair of eyes laterad on the dorsal plate and anteriormost seta (possibly vi) remote from a naso (Figure 2B) is reminiscent of some modern Teneriffiidae, see e.g. Bernardi et al. (2012). The general habitus is similar to some extant Austroteneriffia Womersley, 1935 species (e.g. Khanjani et al. 2014: Figure 1), but the fossil does differ in the leg chaetotaxy and the absence of pectinate claws.
Since the available material in its current state of preservation does not allow a clear resolution of its familial or generic affinities we prefer to treat Sejus bdelloides as a putative Anystina and refer the material to ?Anystoidea incertae sedis. Given that S. bdelloides cannot be meaningfully placed in a known genus – and raising an extinct genus defined on an incomplete character set would be unhelpful – we consider it best to be to formally treat the species name as a nomen dubium. It is not a nomen nudum (sensu Oudemans 1937) as Koch and Berendt’s name is clearly associated with a (brief) description and a figure.
The principal conclusion of our study is that neither Sejida nor Trigynaspida have a demonstrable fossil record. A handful of putative mesostigmatans have been reported from the Cretaceous. Poinar (1998: fig. 1C) figured a specimen from New Jersey amber (90 – 94 Ma), while Peñalver et al. (2017) mentioned their presence in the slightly older Burmese (ca. 99 Ma) and Lebanese (ca. 130 Ma) ambers respectively. However none of these Cretaceous records have been formally described, confirmed as Mesostigmata, or assigned to a higher group. Thus only Monogynaspida is unequivocally represented as fossils; whereby nine of the 77 known families (sensu Beaulieu et al. 2011) have now been recorded (Table 1). Both of the two main monogynaspid clades (i.e. Uropodina and Gamasina) can be traced back to the Eocene (44 – 49 Ma), although this rather young age remains a puzzle given that there are mid-Cretaceous records of two of the other three parasitiform clades, namely Opilioacarida (Dunlop and Bernardi 2014) and Ixodida (ticks) (Poinar and Brown 2003; Poinar and Buckley 2008; Chitimia-Dobler et al. 2017; Peñalver et al. 2017). The final parasitiform group, Holothyrida, lacks a fossil record. By contrast, acariform mites go back over 410 million years to the Early Devonian (Hirst 1923). We hope that Mesozoic ambers will begin to yield further fossil Mesostigmata, and we expect that most of the major parasitiform lineages will prove to be considerably older than their present record indicates. The Late Jurassic origins implied by Peñalver et al. (2017: fig 10) are almost certainly an underestimate, given that most orders of arachnids have Palaeozoic representatives.
We thank Christian Neumann for access to the Berendt collection in Berlin, two anonymous reviewers for valuable comments on a previous version of the typescript and Ekaterina Sidorchuk for editorial comments and assistance with producing one of the figures during a visit to Berlin.
Aoki J. 1974. [On the fossil mites in Mizunami amber from Gifu Prefecture, Central Japan]. Bulletin of the Mizunami Fossil Museum, 1: 397-399 [in Japanese with English summary].
Beaulieu F., Dowling A.P.G, Klompen H., Moraes G.J. de, Walter D.E. 2011. Superorder Parasitiformes Reuter, 1909. In: Zhang Z.-Q. (Ed.). Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa, 3148: 123-128.
Bernardi L.F. de O., Pellegrini T.G., Ferreira R.L. 2012. New species of Neoteneriffiola (Acari: Trombidiformes: Teneriffiidae) from Brazilian caves: geographical distribution and ecological traits. International Journal of Acarology, 38: 410-419. doi:10.1080/01647954.2012.662246 ![]()
Błaszak J., Cokendolpher J.C., Polyak V.J. 1995. Paleozercon cavernicolous, a new genus and new species of fossil mite from a cave in the southwestern U.S.A. (Acari, Gamasida: Zerconidae). International Journal of Acarology, 21: 253-259. doi:10.1080/01647959508684068 ![]()
Chitimia-Dobler L., De Araujo B.C., Ruthensteiner B., Pfeffer T., Dunlop J.A. 2017. Amblyomma birmitum a new species of hard tick in Burmese amber. Parasitolology, 144: 1411-1448. doi:10.1017/S0031182017000853 ![]()
Dabert M., Witalinski W., Kazmierski A., Olszanowski Z., Dabert J. 2010. Molecular phylogeny of acariform mites (Acari, Arachnida): strong conflict between phylogenetic signal and long-branch attraction artifacts. Molecular Phylogenetics and Evolution, 56: 222-241. doi:10.1016/j.ympev.2009.12.020 ![]()
Dunlop J.A., Bernardi L.F. de O. 2014. An opilioacarid mite in Cretaceous Burmese amber. Naturwissenschaften, 101: 759-763. doi:10.1007/s00114-014-1212-0 ![]()
Dunlop J.A., Falkenhagen R. 2014. Raubmilbe in Aragonit. Fossilien, 2014(3): 53-55.
Dunlop J.A., Kontschán J., Zwanzig M. 2013. Fossil mesostigmatid mites (Mesostigmata: Gamasina, Microgyniina, Uropodina), associated with longhorn beetles (Coleoptera: Cerambycidae) in Baltic amber. Naturwissenschaften, 100: 337-334. doi:10.1007/s00114-013-1031-8 ![]()
Dunlop J.A., Kontschán J., Walter D.E., Perrichot V. 2014. An ant associated mesostigmatid mite in Baltic amber. Biology Letters, 10(9): 20140531. doi:10.1098/rsbl.2014.0531 ![]()
Dunlop J.A., Sempf C., Wunderlich J. 2010. A new opilioacarid mite in Baltic amber. In: Nentwig W., Schmidt-Entling M., Kropf C. (eds). European Arachnology, 2008: 59-70.
Giebel C.G. 1856. Die Insekten und Spinnen der Vorwelt mit steter Berücksichtigung der lebenden Insekten und Spinnen; monographisch dargestellt. Leipzig, 511 pp
Giribet G., Vogt L., Pérez González A., Sharma P., Kury A.B. 2009. A multilocus approach to harvestman (Arachnida: Opiliones) phylogeny with emphasis on biogeography and the systematics of Laniatores. Cladistics, 25: 1-30.
Hedin M., Starrett J., Akhter S., Schönhofer A.L., Shultz J.W. 2012. Phylogenomic resolution of Paleozoic divergences in harvestmen (Arachnida, Opiliones) via analysis of next-generation transcriptome data. PloS ONE, 7(8): e42888. doi:10.1371/journal.pone.0042888 ![]()
Hirschmann W. 1971. A fossil mite of the genus Dendrolaelaps (Acarina, Mesostigmata, Digamasellidae) found in amber from Chiapas, Mexico. University of California Publications in Entomology, 63: 69-70.
Hirschmann W., Wisniewski J., Kaczmarek S. 1991. Gangsystematik der Parasitiformes Teil 530. Weltweite Revision der Ganggatttung Sejus C.L. Koch 1836 (Trichopygidina). Neubeschriebung von 26 Sejus-Arten. Wiederbeschriebung der Typenart. Acarologie, 38: 136-214.
Hirst S. 1923. On some arachnid remains from the Old Red Sandstone (Rhynie Chert bed, Aberdeenshire). Annals and Magazine of Natural History, (9)12: 455-474. doi:10.1080/00222932308632963 ![]()
Khanjani M., Hoseini M.A., Asali Fayaz B. 2014. Austroteneriffia kohrrambadiensis n. p. (Acari: Teneriffiidae): a new species from southwestern Iran, with a description of the male. Acarologia, 54: 69-79. doi:10.1051/acarologia/20142116 ![]()
Koch, C.L. 1836. Deutschlands Crustaceeen, Myriapoden und Arachniden, Herausgegeben von Herrich-Schäffer, Regensburg, Fasc. 1.
Koch C.L., Berendt G.C. 1854. Die im Bernstein befindlichen Myriapoden, Arachniden und Apteren der Vorwelt. In: Berendt G.C. (Ed.) Die in Bernstein befindlichen organischen Reste der Vorwelt gesammelt in Verbindung mit mehreren bearbeitetet und herausgegeben 1. Berlin, Nicolai, 124 pp.
Kramer, P. 1877. Grundzüge zur Systematik der Milben. Archiv für Naturgeschichte, 43: 215-247.
Lindquist E.E., Krantz G.W., Walter D.E. 2009. Order Mesostigmata. In: Krantz G.W., Walter D.E. (Eds) A Manual of Acarology. Third Edition. Texas Tech University Press, Lubbock, Texas, pp. 124-231.
Lyubarsky G.Y., Perkovsky E.E. 2012. The first Eocene species of the genus Cryptophagus (Coleoptera, Clavicornia, Cryptophagidae). Vestnik zoologii, 46: 36-40.
Oudemans 1937. Kritisch Historisch Overzicht Der Acarologie, 3de Gedeelte, band C, 1937
Peñalver E., Arillo A., Delclòs X., Peris D., Grimaldi D.A., Anderson S.R., Nascimbene P.C., Pérez-de la Fuente R. 2017. Ticks parasitized feathered dinosaurs as revealed by Cretaceous amber assemblages. Nature Communications, 8: 1924.
Perkovsky E.E., Rasnitsyn A.P., Vlaskin A.P., Taraschuk M.V. 2007. A comparative analysis of the Baltic and Rovno amber arthropod faunas: representative samples. African Invertebrates, 48: 229-245.
Petrunkevitch A.I. 1955. Arachnida. 42-162. In: Moore R.C. (Ed.) Treatise on invertebrate paleontology, Part P, Arthropoda 2. Geological Society of America, Boulder, and University of Kansas Press, Lawrence, xvii + 181 pp.
Poinar Jr G.O. 1998. Fossils explained 22: Palaeontology of amber. Geology Today, July–August 1998: 154-160. doi:10.1046/j.1365-2451.1998.014004154.x ![]()
Poinar Jr G.O., Brown A.E. 2003. A new genus of hard ticks in Cretaceous Burmese amber (Acari: Ixodida: Ixodidae). Systematic Parasitology, 54: 199-205. doi:10.1023/A:1022689325158 ![]()
Poinar Jr G.O., Buckley R. 2008. Compluriscutula vetulum (Acari: Ixodida: Ixodidae), a new genus and species of hard tick from Lower Cretaceous Burmese amber. Proceedings of the Entomological Society of Washington, 110: 445-450. doi:10.4289/07-014.1 ![]()
Ramsay G.W. 1960. Sub-fossil mites from the Hutt Valley. Transactions of the Royal Society of New Zealand, 88: 575-576.
Sayre R.M., Smiley R.L., Walter, D.E. 1992. Report of a teneriffiid mite (Acari) in Baltic amber and notes on recent discoveries. International Journal of Acarology, 18: 303-305. doi:10.1080/01647959208683964 ![]()
Scudder S.H. 1891. Index to the known fossil insects of the world including myriapods and arachnids. Bulletin of the United States Geological Survey, 71: 1-744.
Selden P.A. 1993. Arthropoda (Aglaspidida, Pycnogonida and Chelicerata). Pp. 297-320. In: Benton M.J. (Ed.). The Fossil Record 2. Chapman & Hall, London. Xvii + 845 pp.
Vitzthum H. 1943. Acarina. Bronns Klassen und Ordnung der Tierreichs, Band 5, I Abteilung. Arachnoidea, 1. Lieferung. Akademische Verlagsgesellschaft, Leipzig.
Walter D.E. 2013. A new genus and family of sejine mites (Acari, Parasitiformes, Mesostigmata, Sejoidea) based on new species from Lord Howe Island and Brazil, and a redescription of Sejus americanus (Banks, 1902). Zootaxa 3691: 301-323. doi:10.11646/zootaxa.3691.3.1 ![]()
Walter D.E., Proctor H. C. 1999. Mites. Ecology, Evolution and Behaviour. CABI Publishing, Wallingford.
Walter D.E., Proctor H.C. 2013. Mites: Ecology, Evolution and Behaviour. 2nd Edition – Life at a Microscale. Springer, Amsterdam. 494 pp.
Walter D.E., Lindquist E E., Smith I.M., Cook D.R., Krantz G.W. 2009. Order Trombidiformes. In: Krantz G.W., Walter D.E. (Eds) A Manual of Acarology. Third Edition. Texas Tech University Press, Lubbock, Texas. Pp. 233-420.
Weitschat W., Wichard W. 2010. Baltic amber. In: Penney D. (Ed.) Biodiversity of fossils in amber from the major world deposits. Siri Scientific Press, Manchester, pp. 80-115.
Witaliński W. 2000. Aclerogamasus stenocornis sp. n., a fossil mite from the Baltic amber (Acari: Gamasida: Parasitidae). Genus, 11: 619-626.
Wood H.M., Matzke N.J., Gillespie R.G., Griswold C.E. 2013. Treating fossils as terminal taxa in divergence time estimation reveals ancient vicariance patterns in the palpimanoid spiders. Systematic Biology, 62: 264-284. doi:10.1093/sysbio/sys092 ![]()
Zacharda M., Krivolutsky D.A. 1985. Prostigmatic mites (Acarina: Prostigmata) from the Upper Cretaceous and Paleogene amber of the USSR. Věstník Československé společnosti zoologické, 49: 147-152.



2018-01-30
Date accepted:
2018-04-16
Date published:
2018-07-25
Edited by:
Sidorchuk, Ekaterina

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Dunlop, Jason A.; Walter, David E. and Kontschán, Jenő
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)