



A new species and new record of Gaeolaelaps Evans and Till (Acari: Mesostigmata: Laelapidae) from Iran
Khalesi, Talat1 and Kazemi, Shahrooz2
1Department of Biodiversity, Institute of Science and High Technology and Environmental Sciences, Graduate University of Advanced Technology, Kerman, Iran, P. O. Box: 76315-117.
2✉ Department of Biodiversity, Institute of Science and High Technology and Environmental Sciences, Graduate University of Advanced Technology, Kerman, Iran, P. O. Box: 76315-117.
2018 - Volume: 58 Issue: 3 pages: 628-639
https://doi.org/10.24349/acarologia/20184259ZooBank LSID: 8859D5BD-AD38-4CCC-9194-1C78931F6D36
Keywords
Abstract
The family Laelapidae currently includes 92 known genera and more than 1300 described species and is morphologically and ecologically the most diverse family of Mesostigmata (Beaulieu et al. 2011; Kazemi et al. 2014; Nemati and Gwiazdowicz 2016; Kazemi and Beaulieu 2016). The predatory hypoaspidine genus Gaeolaelaps Evans & Till, 1966 comprises about 100 described species distributed worldwide (Kazemi et al. 2014). Beaulieu (2009) reviewed the concept of this genus and then Kazemi et al. (2014) revised this concept and presented notes on some morphological attributes, particularly idiosomal gland pores and poroids.
To date, 20 species of Gaeolaelaps have been reported from Iran of which nine species (G. jondishapouri Nemati and Kavianpour; G. iranicus Kavianpour and Nemati; G. farajii Nemati and Mohseni; G. orbiculatus Nemati and Mohseni; G. mossadeghi Kavianpour and Nemati; G. khajooii Kazemi et al.; G. ahangarani Kazemi and Beaulieu; G. lenis Vatankhah and Nemati; G. izajiensis Saeidi et al.) were described as new for science (Nemati and Kavianpour 2013; Nemati and Mohseni 2013; Kavianpour et al. 2013; Kazemi et al. 2014; Kavianpour and Nemati 2014; Saeidi et al., 2016; Vatankhah et al. 2016). The Mangrove or Hara Forests of Qeshm in the Persian Gulf, southern Iran, is a protected area that is dominantly covered by Avicennia marina. The mite fauna of Mangrove forests in southern Iran is almost unknown. In this paper we describe a new species of Gaeolaelaps based on adult female and male specimens collected in citrus orchards in Kerman Province, southeastern Iran, and also report G. schusteri (Hirschmann, 1969) for the first time from Hara Forests in northern of the Persian Gulf, Hormozgan Province, Iran.
Mite specimens of the new species were collected in Dosari and Jiroft regions, Kerman Province, southeastern Iran, and specimens of G. schusteri collected in Mangrove forests of Qeshm Island in the Persian Gulf, southern Iran. Mites were extracted from the samples using Berlese-Tullgren funnels, cleared in Nesbitt's fluid and then mounted in Hoyer's medium on microscope slides. Also, the holotype of G. queenslandicus (Womersley, 1956) was examined by junior author (SK).
Morphological observations, measurements and illustrations were made using a compound microscope equipped with differential interference contrast and phase contrast optical systems, and a drawing tube (Olympus BX51). All given measurements are in micrometers (μm). Dorsal shield length was taken from the anterior to posterior shield margins along the midline, and the shield width from the lateral margins at the level of setae r3 and also gland pores gd9; the length and width of the idiosoma, including the dorsal shield and the soft marginal cuticle, were also measured at the same levels (width only at level of setae j6). The width of the sternal shield was taken from lateral margins of the shield at the level of setae st2, and its length from the anterior to posterior margins along the midline. The length of epigynal shield was measured from the anterior margin of the hyaline extension to the posterior margin of the shield along the midline; shield width was taken at level of st5. The anal shield length and width were measured along the midline from the anterior to posterior margins, including the cribrum, and at the broadest point, respectively. The length of male holoventral shield was taken from the anterior to posterior shield margins (including the gonopore) and its width at level of setae st2 and also at broadest point. Legs lengths were measured from the base of the coxae to the apex of tarsi, excluding ambulacra (stalk, claws and pulvillus). The lengths of the second cheliceral segment were taken from their base to the apex of the fixed digit, and their widths at the broadest point. The lengths of the fixed cheliceral digits were measured from the dorsal poroid to the apex and that of the movable digits from the base to apex.
The notation for idiosomal setae follows that of Lindquist & Evans (1965) adapted by Evans & Till (1965, 1966) and Lindquist (1994), and that for leg and palp setae follows that of Evans (1963a, 1963b). The notation for idiosomal pore-like structures as glandular openings (circular symbols) versus poroids (oval-shape symbols) follows mostly Athias-Henriot (1971, 1975), adapted by Kazemi et al. (2014).
Hypoaspis (Gaeolaelaps) Evans & Till, 1966: 159. Type species: Laelaps aculeifer G. Canestrini, 1884, by original designation.
Diagnosis — The genus diagnosis of Beaulieu (2009) and modified by Kazemi et al. (2014) were followed.
ZOOBANK: BA095B1C-5382-47C8-811B-4CC28B554936 ![]()
Diagnosis (adult female and male) — Dorsal shield partly covering dorsal idiosoma, narrowing from level of setae r3 to that of gland pore gd9, then sharply tapering until a rounded posterior end, shield bearing 39 pairs of setae, including px2-3, and seven pairs of gland pores (gd1-2, gd4-5, gd6, gd8-9), shield reticulated posterior to level of setae j6-s5. Adult female sternal shield longer than broad, shield smooth medially, reticulate laterally, gv1 present near posterior margin of shield. Peritremes long, anteriorly reaching to mid-level of coxae I. Palp tarsal claw two-tined. Deutosternal groove with four anterior rows of denticles wider than two posterior rows. Cribrum with anteriormost rows of spicules extending to anterior level of para-anal setae. Male with holoventral shield; spermatodactyl short and apically curved dorsally; male also differs from female by its dorsal shield posteriorly relatively broader than that of female (relative width at level of r3/gd9 in male=1.8, in female=2); internal malae (lateral projections absent).
Description
Adult female (n=4) Figures 1-3, 4A-B
Dorsal idiosoma (Figure 1A, 1D) — Idiosoma length 638 – 685, width 385 – 394. Dorsal shield length 564 – 658, width at level of setae r3 324 – 342 and 159 – 171 at level of gland pore gd9; length of dorsal shield setae: j1 43 – 48, j2 55 – 61, j3 62 – 69, j4-5 56 – 64, j6 44 – 49, z1 46 – 53, z2-5 58 – 64, z6 42 – 47, s1 43 – 48, s2 49 – 55, s3 60 – 68, s4 63 – 72, s5-6 55 – 62, r2 60 – 69, r3 75 – 82, r4-5 51 – 61, J1-3 42 – 49, J4 42 – 54, J5 36 – 42, Z1-4 52 – 67, Z5 64 – 76, S1-4 49 – 64, S5 59 – 69, setae J4-5 and Z5 with few barbs, others smooth (Figure 1D); shield with 23 pairs of pore-like structures, including 16 pairs of poroids and seven pairs of gland pores (gd1-2, gd4-6, gd8-9). Dorsolateral soft integument bearing setae r6 34 – 37, R1-4 29 – 40, R5 43 – 50, R6 50 – 58 and two pairs of UR 30 – 42, and two pairs of poroids, idR3 and one pair of ivo.





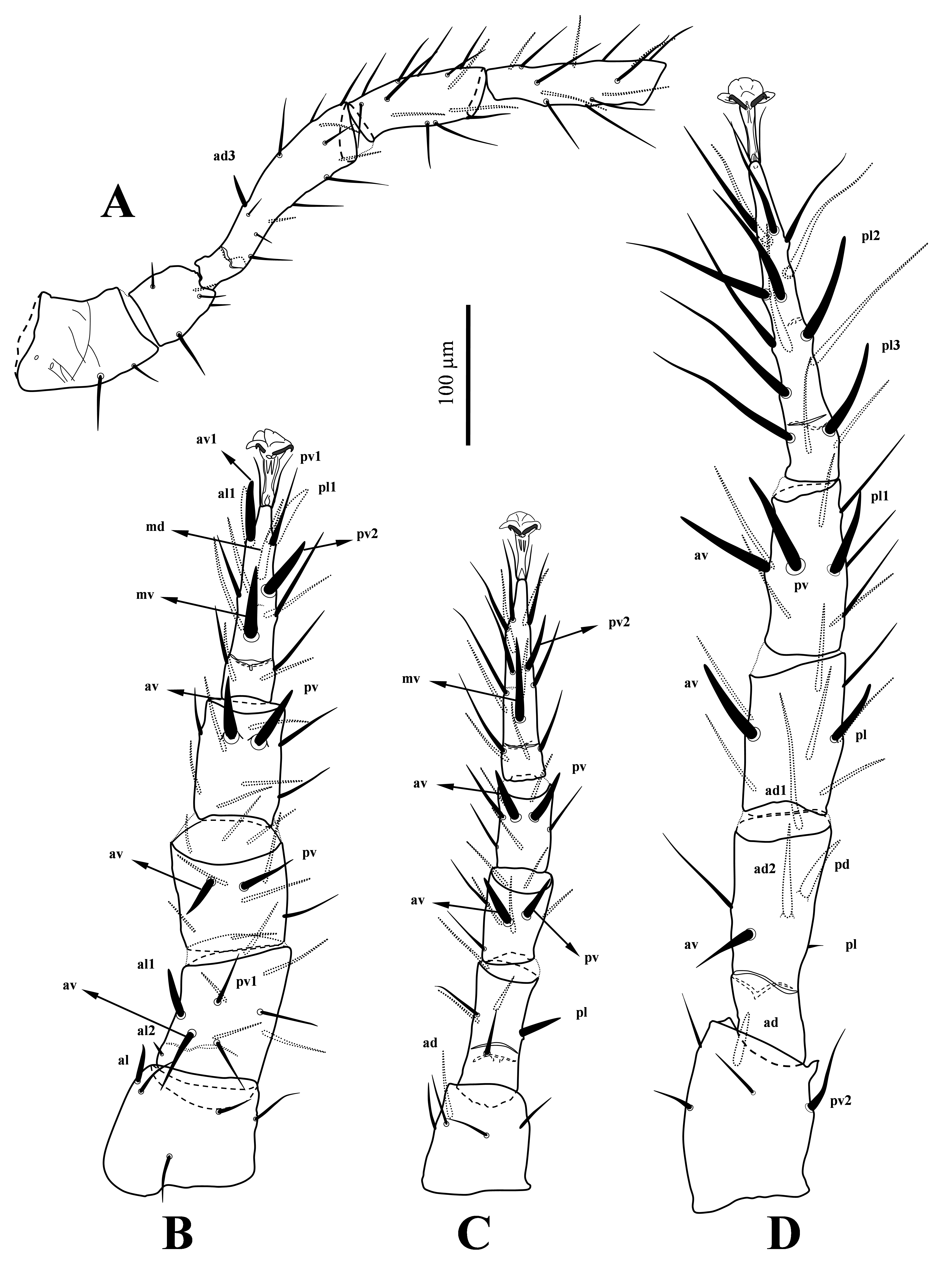
Ventral idiosoma (Figures 2A-B, 4B) — Tritosternum with a narrow columnar base, length 44 – 48, width 14 – 16 at base and 7 – 9 at apex, and two sparsely pilose laciniae, length 99 – 113, fused for 10 – 13 basally. Presternal region weakly sclerotised, lineate and slightly granulate. Sternal shield length 161 – 179, widths at level of st2 and st3 116 – 124 and 137 – 146, respectively, lateral regions of shield reticulate, medially very faintly reticulate, anterior margin of shield slightly concave, posterior margin irregularly straight, anterolateral edges of shield extending between acetabuli I-II bearing gland pores gvb; shield with three pairs of smooth setae: st1 46 – 52, st2 45 – 48, st3 41 – 45 and two pairs of poroids, iv1 slit-like, iv2 round and located midway between setae st1-2, gland pores gv1 inserted near posterior margin of shield. Metasternal setae st4 35 – 40 and associated poroids (iv3) on soft cuticle flanked by free endopodals between coxae III-IV. Epigynal shield almost tongue-shaped, length 181 – 196, width 71 – 73, anterior margin of shield irregularly convex, posterior eight irregular cells flanked by a median inverse Y-shaped ornamentation; setae st5 31 – 36 on lateral margins of shield. Poroids iv5 on soft cuticle, almost at level of tip of st5. A pair of narrow paragenital platelets 24 – 27 long flanked epigynal shield. Anal shield pear-shaped, length 81 – 86, width 61 – 65, anterior region of shield surface with reticulate ornamentation; circumanal setae smooth, post anal seta 40 – 45 longer than para-anals 37 – 40; cribrum well developed, anteriormost row of spicules reaching anterior level of para-anal setae; gland pores gv3 present on lateral margins of shield at level of para-anal setae. Opisthogastric soft integument bearing nine pairs of setae, JV1-4 33 – 43, JV5 59 – 72, ZV1 32 – 36, ZV2-4 36 – 43 and four pairs of poroids (excluding iv5). One pair of narrow platelets 17 – 22 long laterad of ZV1. Metapodal plates divided into two small platelets, primary metapodals (most lateral) larger, narrow, oriented obliquely, and secondary metapodals (more median) minute, oriented transversely. Peritrematal shields narrow, posteriorly reaching mid-level of acetabuli IV, well developed almost at anterior half-length of peritremes, but apically narrowly fused to dorsal shield behind setae z1 level; shields with seven pairs of pore-like structures as shown in Figure 2B. Exopodal platelets between coxae II-III small and subtriangular, those behind coxae III-IV narrow and fused to parapodal platelets posteriorly, gland pores gv2 present on parapodal platelets. Spermatheca as shown in Figure 4B.
Gnathosoma (Figures 1B-C, 2C, 4A) — Anterior margin of epistome densely denticulate (Figure 1B). Corniculi horn-like; internal malae fringed, with one pair of thin contiguous median projections reaching anterior apex of corniculi, and one pair of thicker lateral projections; labrum considerably longer than internal malae; hypostomal and capitular setae smooth: h1 68 – 72, h2 29 – 32, h3 64 – 69, pc 42 – 46, deutosternal groove with six rows of denticles, two basal rows each with 14 – 22, four anterior rows with 29 – 31 denticles and 1.3x as wide as basal rows; rows of denticles flanked anteriorly and posteriorly by a smooth ridge (Figure 2C). Basal segment of chelicerae length 83 – 89; second cheliceral segment length 229 – 241, fixed digit 87 – 94 long, bearing ca. 15 teeth, including gabelzhan; setaceous pilus dentilis small. Movable digit of chelicera length 102 – 109, bidentate. Palp 247 – 252 long, palp chaetotaxy normal for Laelapidae; all setae smooth, palptarsus apotele with two tines (Figure 1C).
Legs (Figures 3A-D) — Leg chaetotaxy normal for Laelapidae (sensu Evans and Till 1965). All legs with ambulacrum, lengths of legs I-IV 684 – 754, 527 – 584, 479 – 515, 761 – 808, respectively. Lengths of femora I 148 – 169, II 94 – 110, III 109 – 116, IV 169 – 187, genua I 94 – 106, II 88 – 96, III 65 – 71, IV 103 – 119; tibiae I 123 – 142, II 85 – 95, III 65 – 71, IV 119 – 132, tarsi I 182 – 199, II 140 – 151, III 144 – 157, IV 212 – 229; pretarsi I 50 – 53, II-IV 52 – 56. Setae on female legs mostly simple and slender, except following setae: al1 on femur II; pv on tibia II; al1, av1, pl1, pv2 on tarsus II; av on genu III; av, pv on tibia III; ad on trochanter IV; pd on femur IV; av, pl on genu IV; av, pv on tibia IV; pl2, pl3 on tarsus IV thickened and spur-like (apically blunt); ad3 on femur I; al on trochanter II; av on femur II; pv2 on trochanter IV; ad1 on femur IV slightly thickened; av, pv on genu II; av on tibia II; md, mv on tarsus II; pl on femur III; pv on genu III; mv, pv2 on tarsus III; ad2, av on femur IV; pl1 on tibia IV thickened and spine-like; setae on tarsus IV mostly elongate and thickened (Figures 3A-D).
Adult male (n=2) Figures 4C-G


Dorsal idiosoma — Idiosoma length 467 – 498, width 276 – 285, dorsal shield length 455 – 484, width at level of setae r3 259 – 272 and 136 – 154 at level of gd9, length of dorsal shield setae: j1 35 – 37, j2 49 – 53, j3 51 – 52, j4 45 – 46, j5 47 – 48, j6 37 – 38, z1 46 – 49, z2 50 – 52, z3 52 – 55, z4 54 – 55, z5 43 – 46, z6 41 – 42, s1 33 – 38, s2 49 – 56, s3 60 – 66, s4 51 – 56, s5 47 – 51, s6 54 – 60, r2 50, r3 62 – 66, r4 46 – 57, r5 48 – 52, r6 33 – 38, J1 39 – 40, J2 35 – 37, J3 35 – 38, J4 39 – 41, J5 38 – 40, Z1 45, Z2 39, Z3 46 – 48, Z4 51 – 55, Z5 58 – 60, S1 52 – 57, S2 40 – 45, S3 49 – 50, S4 48 – 50, S5 54 – 56; other characters of shield as those in female.
Ventral idiosoma (Figure 4C) — Holoventral shield length 407 – 439, widths at level of st2 and ZV1 (broadest point) 100 – 106 and 132 – 163, respectively; shield reticulate throughout, except median area between setae st2-4; with 10 pairs of smooth setae, excluding circumanals: st1-2 35 – 37, st3 33 – 34, st4-5 24 – 27, JV1 27, JV2-3 30 – 31, ZV1 23, ZV2 28 – 30, postanal seta 42 longer than para-anals 29 – 32; bearing four pairs of poroids and two pairs of gland pores (gv1, gv3); gland pores gvb on free, narrow endopodal platelets between acetabuli I-II. Metapodal platelets apparently fused to holoventral shield. Other characters same to those in female.
Gnathosoma (Figures 4D-E) — Internal malae with a pair of median projections slightly longer than corniculi, without lateral projections, and margins fringed (Figure 4D). Length of hypostomal setae: h1 47 – 48, h2 23 – 25, h3 44 – 46, pc 30 – 34. Basal segment of chelicera 38 – 42 long; second segment of chelicera 153 long; cheliceral fixed digit 42 long with five teeth anterior to pilus dentilis, including four small teeth and a larger subapical tooth; movable digit unidentate 59 long, spermatodactyl relatively short, free for 20 – 22 μm. Palps 178 – 185 long. Other characters similar to those in female.
Legs (Figures 4F-G) — Lengths of legs I-IV 553 – 612, 435 – 454, 410 – 425, 617 – 660, respectively. Lengths of femora I 112 – 115, II 70 – 78, III 87 – 89, IV 130 – 134; genua I 65 – 68, II 58 – 71, III 44 – 52, IV 77 – 80; tibiae I 92 – 94, II 55 – 71, III 46 – 52, IV 86 – 94; tarsi I 140 – 146, II 101 – 115, III 113 – 117, IV 167 – 181 and their pretarsi I 47 – 49, II 44 – 47, III 38 – 43, IV 45 – 47. Leg chaetotaxy and shape of setae mostly similar to those in female, except setae av and pv1 on femur II (av and pv1 thickened in male vs. av in female only slightly thickened, pv slender, and both setae shorter than those in female), and setae av and pv on genu III (av two times longer than pv in male vs. av only slightly longer than pv in female).
Remarks — Gaeolaelaps mirzakhaniae n. sp. can be readily distinguished from all other species of the genus by the combination of the following characters: (1) dorsal shield narrowed behind level of setae r3, then abruptly tapered past gland pores gd9 to end into a round, V-shaped apex; (2) dorsal shield bearing 39 pairs of setae, including px2-3, and seven pairs of gland pores, including gd4; (3) palptarsus apotele two tined. The shape of the dorsal shield of G. mirzakhaniae n. sp. is a characteristic attribute for the species and only G. queenslandicus (Womersley, 1956) has a similar dorsal shield, but these two species can be easily separated by several differences such as (1) palptarsus apotele two tined vs. three tined in G. queenslandicus; (2) deutosternal groove with four anterior denticulate rows wider than two posterior rows vs. three anterior denticulate rows wider than three posterior rows in G. queenslandicus; (3) presence of 39 pairs of setae on dorsal shield vs. 37 pairs in G. queenslandicus (px2-3 absent, rarely px3 present). Also, in G. mirzakhaniae n. sp. poroids idR3 have been located between setae R2-3, while in other Gaeolaelaps species they inserted in usual region of these poroids, between R3-4 (Beaulieu 2009; Kazemi et al. 2014).
Material examined — Holotype female: Southeastern Iran, Kerman Province, Dosari Region (28°25´36˝N; 57°57´19˝E), altitude 640 m above sea level, 19 August 2016, collected in soil and litter of a citrus orchard, coll. T. Khalesi, deposited in Acarological Collection, Institute of Science and High Technology and Environmental Sciences, Graduate University of Advanced Technology, Kerman, Iran (ACICST). Paratypes: three females and one male with same data as holotype (ACICST), one female with same data as holotype, will be deposited in Jalal Afshar Museum of Zoology, Tehran University, Karaj, Iran (JAMZ), one female and one male in another citrus orchard of the same region (28°25´55˝N; 57°57´26˝E), altitude 644 m above sea level, 22 April 2016, coll. T. Khalesi (ACICST), three females in Kerman Province, Jiroft Region, Dolat-Abad, (28°36´37˝N; 57°48´10˝E), altitude 637 m above sea level, 23 Sept. 2017, collected in soil of a citrus orchard, coll. F. Dadollahi (ACICST).
Etymology — The species is named in honor of Professor Maryam Mirzakhani (3 May 1977 – 15 July 2017) who was an Iranian mathematician that became both the first woman and the first Iranian honored with the “Fields Medal" in 2014, the most prestigious award in mathematics.
Hypoaspis schusteri Hirschmann, 1966: 26.
Hypoaspis schusteri Costa, 1974: 221.
Gaeolaelaps schusteri Beaulieu, 2009: 36; Kazemi et al., 2014: 504; Trach, 2016: 10.
Note — In a survey on the mesostigmatic mite fauna of Mangrove forests in Qeshm region, northern of the Persian Gulf, some specimens of G. schusteri (Hirschmann, 1966) were collected and identified. This represents the first report of the species from Iran.
Material examined — Seven females, two males, two deutonymphs and two protonymphs, Southeastern Iran, northern of the Persian Gulf, Hormozgan Province, Qeshm Region (26°52´3.6˝N; 55°35´0.6˝E), 4 May 2017, collected in soil and crashed cockleshells, coll. S. Kazemi (deposited in ACICST); one female with same data (deposited in JAMZ); one female, one male, one deutonymph, Southeastern Iran, Hormozgan Province, northern of the Persian Gulf, Qeshm Region (26°55´27.54˝N; 55°43´42.9˝E), 29 Dec 2017, collected in soil and crashed cockleshells, coll. S. Kazemi (deposited in ACICST).


Womersley (1956) described a new laelapid mite species under the genus Androlaelaps Berlese, A. queenslandicus, based on one female specimen collected in leaf debris in Queensland, Australia. Ryke (1963) transferred this species to the genus Gaeolaelaps, redescribed it based on specimens collected in South Africa and described its male for the first time. Costa (1966) redescribed again the species and additionally described its deutonymph. However, based on examination of some specimens of the species collected in different regions in Iran and the type species, we found some discrepancies between our materials and the previous description/redescriptions including: (1) peritremes reach to level of setae s1 (Figure 5C), but Womersley mentioned to long peritremes only, and Ryke and Costa referred to peritremes reaching to level of coxae I, and reaching to behind setae z1, respectively; (2) dorsal shield has slightly (Figure 5A, and Figure 32 in Ryke (1963)) or distinct (Figure 1 in Costa (1966)) depressions laterally between setae S3-4, but in the original description Figure 48B almost no depressions were illustrated; (3) anterolateral edges of the sternal shield well-developed between coxae I-II, bearing gland pores gvb, and also gv1 in posterior region of the shield (Figures 5B), but without anterolateral edges, gvb and gv1 in the original description and also redescriptions; (4) a pair of narrow parapodal plates bearing gland pores gv2 and also small exopodal platelets between coxae III-IV present, not mentioned in the original description and redescriptions. Also, anal shield is considerably longer than wide, with ratio of length/width 1.6-1.7 (Figures 5D).
Previously, Kazemi et al. (2014), who examined a single poorly mounted specimen of G. queenslandicus, stated that gland pores gd4 were not present in G. queenslandicus. However, a recent examination showed that gd4 are present in G. queenslandicus. Also, this species has gland pores gv1 near the posterior margin of sternal shield which may represent a species group within Gaeolaelaps. These glands or their remnants are also present in some other Gaeolaelaps species, including G. aculeifer (G. Canestrini), G. angusta (Karg) (probably junior synonym of G. queenslandicus), G. nolli (Karg), G. kargi (Costa), G. khajooii, G. ahangarani, G. jondishapouri, G. cf farajii and the newly described species, G. mirzakhaniae.
We wish to thank Owen Seeman (Queensland Museum, Brisbane, Australia) and two anonymous reviewers for their valuable suggestions, Farshid Dadollahi for his help with collecting some specimens of the new species, and Peter Hudson (South Australian Museum) for his help to loan the type species of Gaeolaelaps queenslandicus. This work was partly supported by INSF, under grant number 96000698.
Athias-Henriot C. 1971. La divergence néotaxique des Gamasides (Arachnides). Bull. Sc. Bourgogne, 28: 93-106.
Athias-Henriot C. 1975. Nouvelles notes sur les Amblyseiini. 2. Le relevé organotaxique de la face dorsale adulte (gamasides, protoadéniques, Phytoseiidae). Acarologia, 17: 20-29.
Beaulieu F. 2009. Review of the mite genus Gaeolaelaps Evans &Till (Acari: Laelapidae) and description of a new species from North America, G. gillespiei. Zootaxa, 2158: 33-49.
Beaulieu F., Dowling A.P.G, Klompen H., de Moraes G.J., Walter D.E. 2011. Superorder Parasitiformes Reuter, 1909. In: Zhang, Z.-Q. (Ed.). Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa, 3148: 123-128.
Canestrini G. 1884. Acari nuovi o poco noti. Atti del Reale Istituto Veneto di Scienze, Lettere ed Arti (Ser. 6), 2: 693-724 + Plates VI-IX.
Costa M. 1966. A redescription of Hypoaspis queenslandicus (Womersley, 1956) comb. nov. (Acari, Mesostigmata) with notes on the genus Hypoaspis Canestrini. Isr. J. Zool., 15: 141-147.
Costa M. 1966. Mesostigmatic mites (Acari: Mesostigmata) from the Mediterranean shores of Israel. Isr. J. Entomol., 11: 219-228.
Evans G.O. 1963a. Observation on the chaetotaxy of the legs in the free-living Gamasina (Acari: Mesostigmata). Bull. Br. Mus. (Nat. Hist.), Zool., 10: 277-303. doi:10.5962/bhl.part.20528 ![]()
Evans G.O. 1963b. Some observations on the chaetotaxy of the pedipalps in the Mesostigmata (Acari). Ann. Mag. Nat. Hist. (Ser. 13), 6: 513-527. doi:10.1080/00222936308651393 ![]()
Evans G.O., Till W.M. 1965. Studies on the British Dermanyssidae (Acari: Mesostigmata). Part 1. External morphology. Bull. Br. Mus. (Nat. Hist.), Zool., 13: 247-294. doi:10.5962/bhl.part.16752 ![]()
Evans G.O., Till W.M. 1966. Studies on the British Dermanyssidae. 2. Classification. Bull. Br. Mus. (Nat. Hist.), Zool., 14: 107-370.
Hirschmann W. 1966. Gangsystematik der Parasitiformes. Teil 15. Gänge von Litoralmilben und neue Litoralmilbenarten. Acarol. Schrift. Vergl. Milbenk., 11: 219-228.
Karg W. 1965. Larvalsystematische und phylogenetische Untersuchung sowie Revision des Systems der Gamasina Leach, 1915 (Acarina, Parasitiformes). Mitt. Zool. Mus. Ber., 41: 193-340. doi:10.1002/mmnz.19650410207 ![]()
Kavianpour M., Nemati A., Gwiazdowicz D.J., Kocheili F. 2013. A new species of the genus Gaeolaelaps (Acari, Mesostigmata, Laelapidae) from Iran. Zookeys, 277: 1-11. doi:10.3897/zookeys.277.4741 ![]()
Kavianpour M., Nemati A. 2014. Gaeolaelaps (Acari: Laelapidae) mites from Iran with description of a new species. J. Crop Prot., 3: 315-325.
Kazemi S., Beaulieu F. 2016. A new genus and species of Laelapidae (Acari: Mesostigmata) from Iran. Zootaxa, 4200(4): 487-500. doi:10.11646/zootaxa.4200.4.2 ![]()
Kazemi S., Rajaei A., Beaulieu F. 2014. Two new species of Gaeolaelaps (Acari: Mesostigmata: Laelapidae) from Iran, with a revised generic concept and notes on significant morphological characters in the genus. Zootaxa, 3861(6): 501-30. doi:10.11646/zootaxa.3861.6.1 ![]()
Lindquist E.E. 1994. Some observations on the chaetotaxy of the caudal body region of gamasine mites (Acari: Mesostigmata), with a modified notation for some ventrolateral body setae. Acarologia, 35: 323-326.
Lindquist E.E., Evans G.O. 1965. Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina (Acarina: Mesostigmata). Mem. Entomol. Soc. Canada, 47: 1-64. doi:10.4039/entm9747fv ![]()
Nemati A., Kavianpour M. 2013. A new species of Laelapidae (Acari: Mesostigmata) from Iran. J. Crop Prot., 2(1): 63-73.
Nemati A., Mohseni M. 2013. Two new species of Gaeolaelaps (Acari: Laelapidae) from Iran. Zootaxa, 3750(1): 71-82. doi:10.11646/zootaxa.3750.1.5 ![]()
Nemati A., Gwiazdowicz D.J. 2016. A new genus and species of Laelapidae from Iran with notes on Gymnolaelaps Berlese and Laelaspisella Marais & Loots (Acari, Mesostigmata). ZooKeys, 549: 23-49. doi:10.3897/zookeys.549.6891 ![]()
Ryke P.A.J. 1963. Some free-living Hypoaspidinae (Acari : Mesostigmata) from South Africa. Rev. Biol., 5: 1-15.
Saeidi Z., Nemati A., Khalili-Moghadam A. 2016. Description of a new species of Gaeolaelaps (Acari: Laelapidae) from Iran. Zookeys, 612: 31-40. doi:10.3897/zookeys.612.9678 ![]()
Trach V.A. 2016. Three new unusual beetle associated species of the genus Gaeolaelaps (Acari, Mesostigmata, Laelapidae) from Ukraine. Vestnik Zoologii, 50(1): 3-16. doi:10.1515/vzoo-2016-0001 ![]()
Vatankhah F., Nemati A., Esfandiari M., Shishehbor P. 2016. Description of a new species of Gaeolaelaps (Acari: Laelapidae) from Iran, with a key to world species of the genus with short peritremes. Zootaxa, 4121(5): 566-574. doi:10.11646/zootaxa.4121.5.6 ![]()
Womersley H. 1956. On some new Acarina-Mesostigmata from Australia, New Zealand and New Guinea. J. Linn. Soc. Lond. (Zool.), 42: 505-599.



2017-07-26
Date accepted:
2018-02-23
Date published:
2018-06-01
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Khalesi, Talat and Kazemi, Shahrooz
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)