



A contribution to the knowledge of Quadraseta brasiliensis Goff and Gettinger, 1989 (Trombidiformes: Trombiculidae), with description of the deutonymph instar
Jacinavicius, F. C.1 ; Bassini-Silva, R.2 ; Mendoza-Roldan, J. A.3 ; Muñoz-Leal, S.4 ; Hingst-Zaher, E.5 ; Ochoa, R.6 ; Bauchan, G. R.7 and Barros-Battesti, D. M.8
1Laboratório Especial de Coleções Zoológicas, Instituto Butantan, São Paulo, SP; Brazil. Departamento de Medicina Veterinária Preventiva e Saúde Animal, FMVZ-USP, São Paulo, SP, Brazil.
2Laboratório Especial de Coleções Zoológicas, Instituto Butantan, São Paulo, SP; Brazil. Departamento de Medicina Veterinária Preventiva e Saúde Animal, FMVZ-USP, São Paulo, SP, Brazil.
3Laboratório Especial de Coleções Zoológicas, Instituto Butantan, São Paulo, SP; Brazil. Departamento de Medicina Veterinária Preventiva e Saúde Animal, FMVZ-USP, São Paulo, SP, Brazil.
4Departamento de Medicina Veterinária Preventiva e Saúde Animal, FMVZ-USP, São Paulo, SP, Brazil.
5Museu Biológico do Instituto Butantan, Instituto Butantan, São Paulo, SP, Brazil.
6Systematic Entomology Laboratory, United States Department of Agriculture, Agricultural Research Service, Beltsville, MD, USA.
7Electron and Confocal Microscopy Unit, United States Department of Agriculture, Agriculture Research Service, Beltsville, MD, USA.
8✉ Departamento de Medicina Veterinária Preventiva e Saúde Animal, FMVZ-USP, São Paulo, SP, Brazil. Departamento de Patologia Veterinária, Faculdade de Ciências Agrárias e Veterinárias-UNESP, Jaboticabal, SP, Brazil.
2018 - Volume: 58 Issue: 2 pages: 442-456
https://doi.org/10.24349/acarologia/20184252ZooBank LSID: 6AA1C791-3928-4B69-BF75-54EFAF145F24
Keywords
Abstract
Around 3,000 species of chigger mites (Trombidiformes: Trombiculidae) have been described worldwide (Goff et al. 1982), and most of this diversity is known only by the type specimens. The life cycle of chiggers includes seven stages, namely egg, pre-larva, larva, protonymph, deutonymph, tritonymph and adult, yet active stages are larvae, deutonymphs and adults. While the majority of larvae from this group of mites are ectoparasites that feed on terrestrial vertebrates, other active stages are arthropod predators. Because of their parasitism in larval stage, the taxonomy of these mites has been based on this stage (Miyajima and Okumura 1917). The latest revision of chigger genera in the Neotropical and Nearctic regions was published in the 1970’s accounting more than 100 genera, with 19 of these taxa documented for Brazil (Brennan and Goff 1977).
In the Neotropical region, the genus Quadraseta Brennan, 1970 is known only from larval stage and consists in 14 ectoparasitic species associated with small-mammals; however one species, Quadraseta flochi (Brennan and Jones, 1960), can also parasitize birds. Of these Neotropical representatives, Quadraseta brasiliensis Goff and Gettinger, 1989 and Quadraseta pazca (Brennan and Jones, 1964) have been reported from Brazil.
Quadraseta brasiliensis was described from larvae collected on the rodent Hylaeamys megacephalus (Fisher), cited as Oryzomys capito (Olfers), and from the marsupials Gracilinanus agilis (Burmeister), cited as Marmosa agilis, and Monodephis americana (Müller), all captured at an average of 25 km from Brasília (Federal District). In the original description of Q. brasiliensis, Goff and Gettinger (1989) designated the holotype based on larva collected from H. megacephalus, deposited in the MZUSP – Museu de Zoologia da Universidade de São Paulo, São Paulo, Brazil, and paratypes deposited in following three collections: BPBM – Bernice Pauahi Bishop Museum, Honolulu, Hawaii, USA (currently housed in the NMNH collection), OMNH – Sam Noble Oklahoma Museum of Natural History, Norman, Oklahoma, USA, and NMNH (earlier acronym USNM) – United States National Museum of Natural History, Smithsonian Institution, at United States Department of Agriculture, Beltsville, Maryland, USA. Unfortunately no type specimens of Q. brasiliensis were found while examining these collections, however, we found specimens mounted on slides, collected from the same host species and locality, but different date, that allowed us to confirm the identity of our material. Here we redescribe the larva, describe the deutonymph instar obtained from field-collected larvae, report new hosts and localities, and include genetic data for this species in Brazil.
Slide-mounted larvae of Quadraseta deposited in the NMNH, which are housed at the Systematic Entomology Laboratory (BARC-USDA-ARS), and in the Acari Collection from Butantan Institute, São Paulo, Brazil (IBSP), were examined and identified based on the original descriptions of the 14 known Quadraseta species.
Measurements and drawings were made using a Leica DFC 500 digital camera coupled to a Leica DM4000B optical microscope. Extended focal range images were composed using the Leica Application Suite version 2.5.0. Optical micrographs were prepared using Adobe Photoshop v. 13.0 and Inkscape V. 2. All measurements were made in micrometers (μm), followed by maximum, minimum, mean and standard deviation (SD). Micrographs of larvae and deutonymphs were made by the Low Temperature Scanning Electron Microscopy (LT-SEM) at the US Department of Agriculture, Electron and Confocal Microscopy Unit, Beltsville, Maryland as described in Dowling et al. (2010) and Bolton et al. (2014).
We followed the terminology of Goff et al. (1982), with adaptations proposed by Stekolnikov (2008) and Stekolnikov and Daniel (2012) concerning general nomenclature of larval stages. To standardize the nomenclature of the specialized setae of the Prostigmata group, we adopted the terminology used by Wohltmann (2006, 2007) and Kethley 1990. For the deutonymphs, we used the terminology proposed by Audy (1954) and Crossley (1960).
Chigger larvae collected on rodents and marsupials from the municipality of Morro Grande, Cotia, São Paulo were reared until deutonymph stage under laboratory conditions. To achieve this, part of these larvae were placed in Petri dishes, with a mixture of plaster of Paris and activated charcoal with different proportions (3:1, 2:1 and 1:1). The Petri dishes were kept in darkness at room temperature (range 26°C ± 1ºC), and the substrate was daily moistened with distilled water to provide (98% ± 1%) humidity. Once deutonymphs emerged, corresponding skins left on the dish surface were mounted on slides in Hoyer's medium for identification following Walter and Krantz (2009). Eggs of Onychiurus sp. (Insecta: Collembola) and Aedes aegypti (Insecta: Diptera) were offered as food (Lipovsky 1954). One part of the field-collected larvae and laboratory-reared deutonymphs was preserved in 100% ethanol for molecular analyses. Old larval skins and the rest of the material were deposited as voucher at the IBSP collection.
DNA extraction using the Guanidine Isothiocyanate lysis protocol (Chomkzynski 1993) was individually performed to 19 larvae (IBSP12557B) collected from Akodon montensis and to 5 deutonymphs reared from larvae hosted by the same rodent. Each mite was placed in an Eppendorf microtube, and punctured in the idiosomal region with a sterile needle (1.20 * 40 – 18G). After the DNA extraction all exuviae were recovered and mounted in slides, as a voucher.
A PCR targeting a ≈ 500-pb fragment of the 18S ribosomal RNA gene was performed using primers Mite18S-1F (3’-ATATTGGAGGGCAAGTCTGG-5’) and Mite18S-1R (3’-TGGCATCGTTTATGGTTAG-5’) as described by Otto and Wilson (2001). For the gene COI we used the following primers targeting a ≈ 400-600-pb: bcdF01 (CATTTTCHACTAAYCATAARGATATTGG) and bcdR04 (TATAAACYTCDGGATGNCCAAAAAA) as described by Dabert (2008, 2010) with adaptations of Moniuszko et al. (2015); and 772 (TGATTTTTTGGTCACCCAGAAG) and 773 (TACAGCTCCTATAGATAAAAC) as described by Navajas (1994). For each reaction, negative (Milli-Q water free of DNA) and positive controls (chigger mite pools) were used. All PCRs were performed in a Mastercycler Gradient (Eppendorf® California, USA). PCR products with concentrations higher than 20 ng/µl, were selected and purified with ExoSap-IT (GE Healthcare Pittsburgh, PA). Sanger sequencing reactions of the samples were performed in the “Centro de pesquisa sobre Genoma Humano e Células Tronco do Instituto de Biociências da USP”. Obtained sequences were assembled with Sequencing Analysis 5.3.1 software, and submitted to BLAST analyses (Altschul et al. 1990) in order to infer similarities with other mites available in GenBank. Different haplotypes were visually discriminated after an alignment using CLUSTAL W algorithm (Thompson et al. 1994) implemented in Geneious R9 (Kearse et al. 2012).
Quadraseta brasiliensis Goff and Gettinger, 1989: 557.
Diagnosis — Larva. (Figures 1, 2 and 3A; Table 1) Palpal tibia setae branched, galeal setae nude, claviform trichobothria, 2 humeral setae, the first row of idiosomal dorsal setae are arranged in line with 8 setae, genu of leg I with 3 solenidia, the base of microseta is proximal to the base of solenidion on the tarsus of leg I.








Deutonymph. (Figures 4 and 5, Table 2) Sternal area with five branched setae on one side and four branched setae on the other side between coxae I; 8 distinct but minute dorsal teeth in cheliceral each blade, palp tibia with 5 setae branched and 1 dorsal nude seta; base of tibial claw flanked by 2 spoon-shaped accessory setae, palp tarsus with 7 branched setae, 2 nude apical sensory setae, a leaf-shaped tectum, with a single tectal seta, parascutal setae number approximately six on either side of crista, typically arranged at 2-4; three pairs of branched epivalval setae and six pairs of branched centrovalval setae.






Redescription — Larva. Yellowish-white colored when alive. Gnathosoma (Figure 1A and B, 2C) – palpal setal formula B/B/BBB/4B, B = branched seta on the femur; B = branched seta on genu; BBB = branched dorsal, ventral setae, and lateral tibial setae, respectively 4B = four branched with ω on tarsus; odontus trifurcate; cheliceral blade with tricuspid cap; gnathobase punctate, galeal nude setae. Idiosoma – eyes 2/2; anterior larger, on ocular plate, scutum with 1 pair of AL, 1 pair of PL and a single AM seta, PL > AM > AL; 1 pair of claviform trichobothria, covered with setules; scutum with concave lateral margins, posterior margin with two smooth evaginations and anterior margin with two invaginations both turned to the base of trichobothria, as figure 1C, 2E and F. Ninety-four idiosomal setae (material range 84-97 setae), including 1 pair of humeral, 1st row of dorsal idiosomal setae with 8 setae, 2nd row with 8 setae, 3rd row with 8 setae, 4th row with 11 setae and 5th row with 13 setae, totaling 48 dorsal setae (material range 39-48 setae), 2 pairs of sternal, and 26 preanals and 14 postanals, totalizing 40 ventral setae (material range 40-47 setae) (Figures 1H and G, 2D). Legs – seven-segmented, femur divided in basifemur and telofemur, with a pair of claws and a clawlike empodium, onychotriches absent, coxae not striate. Leg I – coxa with 1 branched seta (1B); trochanter 1B; basifemur 1B; telofemur 5B; genu 4B, σ 3, with κ; tibia 8B, Φ 2, with κ; tarsus 21B, with ω, ε, dorsal eupathid (ζ) with a companion seta (z) and subterminal eupathid (ζ), base of ε proximal to base of ω (Figure 1D, 2A and B). Leg II – coxa 1B; trochanter 1B; basifemur 2B; telofemur 4B; genu 3B, σ; tibia 6B, Φ 2; tarsus 16B, with ω, ε, and subterminal eupathid, base of ε proximal to base of ω (Figure 1E). Leg III – coxa 1B on anterior margin, trochanter 1B; basifemur 2B; telofemur 3B; genu 3B, σ, tibia 6B, Φ; tarsus 15B (Figure 1F). The morphometric data are shown in Table 1.
Description — Deutonymph. Dorsal idiosoma – color whitish yellow, eyes absent; The entire body with branched setae on plates, shown in Figure 4B; a leaf-shaped tectum, with a single tectal seta; six parascutal setae on either side of crista, typically arranged at 2-4 (Figure 4C); trichobothria long and slender, proximal portion with numerous short branches, and the distal portion with numerous long thin branches (Figure 4C and E). Ventral idiosoma – Sternal area: with five branched setae on one side and four branched setae on the other side between coxae; six pairs of branched epivalval setae, 2 pairs of oval acetabula present, posterior acetabulum smaller than anterior and three pairs of branched centrovalval setae (Figure 4D); a pair of anal plates, each one with eight branched setae (Figure 4D). Gnathosoma – Cheliceral blade straight, V-shaped, with 8 distinct but minute dorsal teeth, along each. Entire hypostome not divided into lobes, with 4 pairs of nude apical setae; palp femur with 8 branched setae; palp genu with 7 branched setae; palp tibia with 5 setae branched and 1 dorsal nude seta; base of tibial claw flanked by 2 spoon-shaped accessory setae; palp tarsus with 8 branched setae, 2 nude apical sensory setae (ζ) and 1 basal sensory rod (ω) (Figure 4F). Legs – seven-segmented, femur divided in basifemur and telofemur, tarsi with a pair of claws, onychotriches absent; leg I and IV longer than legs II and III; tarsus I oblong in shape, inflated, always longer than tibia I (Figure 4A). All leg segments densely covered with normal setulated setae and the specialized setae present on leg I (tFe–Ta), II (tFe-Ta), III (tFe-Ta), IV (Ge–Ta) (Figure 5A-F). The morphometric data are shown in Table 2.
Remarks — The species Q. brasiliensis resembles the species Q. pazca and Q. mackenziei (Yunker and Brennan, 1964), but is has the first row of dorsal idiosomal setae in lines and the all palpal tibia setae branched. The species Q. brasiliensis differs from the other species in presents 8 setae in the first row of idiosomal dorsal setae, while the other two species have 10 setae. Q. brasiliensis and Q. mackenziei has galeal nude setae, while Q. pazca has galeal branched setae. The trichobothria in Q. brasiliensis and Q. pazca are clavate, while in Q. mackenziei are globose.
Biology — In laboratory conditions, engorged larvae, recently collected from the hosts, kept in Petri dishes with substrate (1:1), remained active for 1-3 days. After this period the larvae enter in the process of quiescence (protonymph), at this stage there is an extrusion of the legs. This process lasted 7-11 days and a deutonymph emerged from the anterior dorsal portion of protonymph, the deutonymphs remained alive for up to 20 days without feeding.
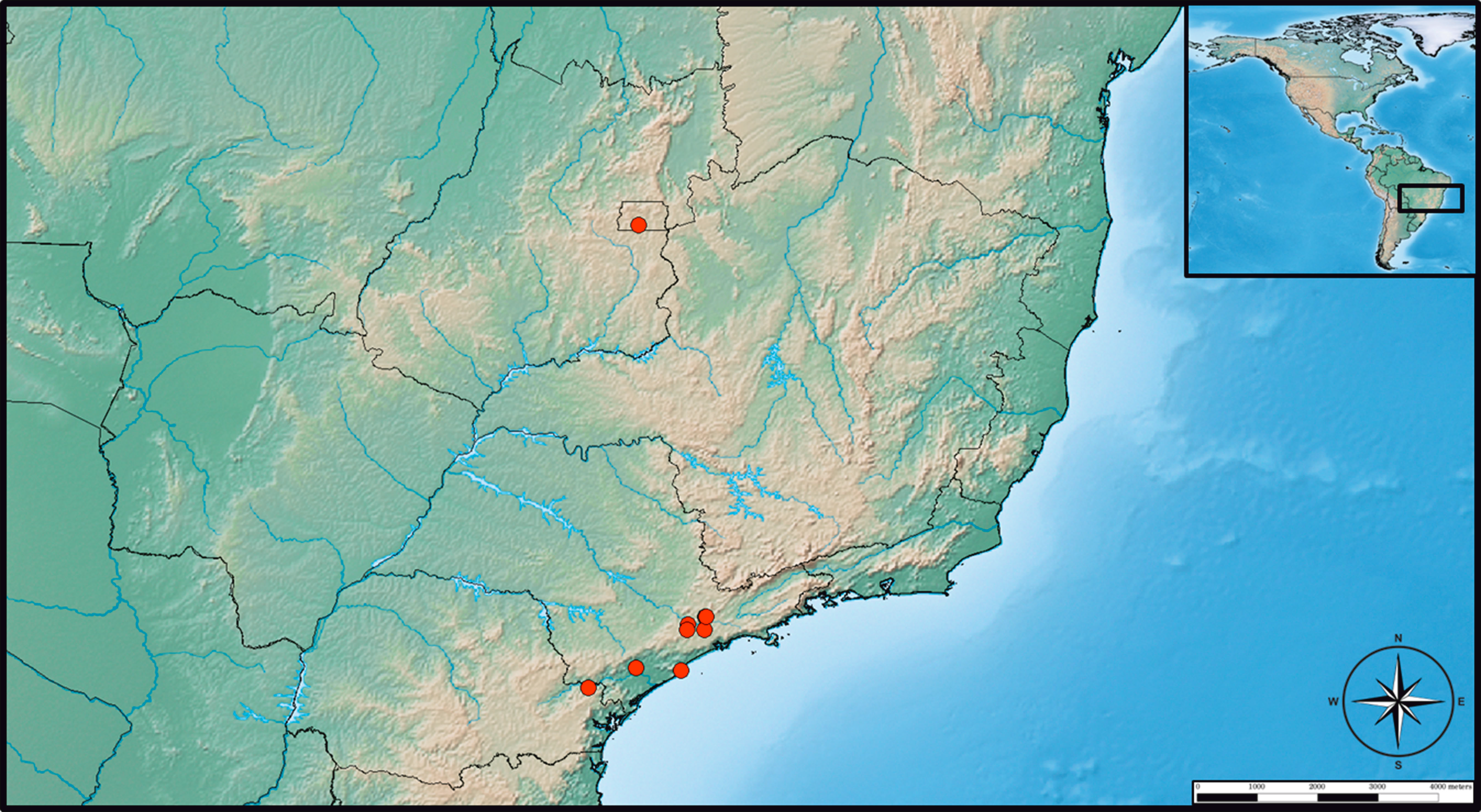
New records (Figure 6) — BRAZIL – 6 larvae, Fazenda Água Limpa (FAL), Brasilia – Federal District, (15°56′45′′S, 47°56′8′′W), 21-V-1984, H. megacephalus (DG494); 10 larvae, same locality, 24-V-1984, Monodelphis americana (DG498); 10 larvae, same locality and host (DG515), 16-VI-1984; 10 larvae, same locality, 21-II-1984, Monodelphis sp. (DG260); 10 larvae, same locality, 22-IV-1984, H. megacephalus (DG394); 10, same locality, 24-II-1984, H. megacephalus (102); 10 larvae, Reserva Ecológica do Instituto Brasileiro de Geografia e Estatística (IBGE) (15°56′41′′S, 47°53′07′′W), 8-VII-1984, no host information (37); 1 larva (IBSP 1154), Serra da Cantareira, São Paulo, state of São Paulo (23°22′44′′S, 46°31′38′′W), 8-IX-1937, Cuniculus paca, Navas J. coll.; 8 larvae (IBSP 10390), Adrianópolis, state of Paraná (24°39′25′′S, 48°59′27′′W) 12-IX-2009, Euryoryzomys russatus; 1 larva (IBSP10521A), Zoológico de São Paulo, São Paulo, state of São Paulo (23°38′57.49′′S, 46°37′19.76′′W), 16-IV-2010, Akodon sp.; 1 larva (IBSP 10522), Barra do Una, Peruíbe, state of São Paulo (24°19′12′′S, 46°59′52′′W), 8-XI-2010, Euryoryzomys russatus; 2 larvae (IBSP 10524A), same locality and host, 18-VI-2010; 1 larva (IBSP 10525A), same data; 2 larvae (IBSP 10526A), same locality and host, 16-VI-2010; 4 larvae (IBSP 10527A), same data; 1 larva (IBSP 10528), same locality and host, 7-XI-2010; 2 larvae (IBSP 10529), same locality and host, 21-IV-2010; 2 larvae (IBSP 10530), same locality and host, 20-IV-2010; 2 larvae (IBSP 10531), same locality and host, 21-IV-2010; 3 larvae (IBSP 10532), same data; 3 larvae (IBSP 10533A), same locality and host, 19-IV-2010; 5 larvae (IBSP 10535A), Serra da Cantareira, São Paulo, state of São Paulo, same host, 12-IV-2011; 2 larvae (IBSP10536), same locality and host, 14-IV-2011; 10 larvas (IBSP 10537), same data; 3 larvae (IBSP10538), same locality and host, 18-XII-2010; 4 larvae (IBSP 10540), same locality and host, 17-II-2010; 1 larva (IBSP 10590D), Condomínio Vila Verde, Itapevi, state of São Paulo, (23°32′56′′S, 46°56′02′′W), 12to16-XII-2011, Didelphis aurita; 6 larvae (IBSP 11096G), Morro Grande, Cotia, state of São Paulo (23°38′58.12′′, 46°57′45.99′′W), 20-VI-2012, Euryoryzomys russatus; 9 larvae (IBSP 11116B), Condomínio Vila Verde, Itapevi, state of São Paulo, 29-XI-2012, Monodelphis sp.; 1 larva (IBSP 11130), Morro Grande, Cotia, state of São Paulo, 22-VII-2012, Monodelphis sp.; 2 larvae (IBSP11131), same locality, 19-X-2012, Euryoryzomys russatus; 1 larva (IBSP 11132), same locality, 23-VII-2012, Thaptomys nigrita; 1 larva (IBSP 11178C), same locality, Akodon montensis; 4 larvae (IBSP 11259C), same locality, 18-IV-2013, Euryoryzomys russatus; 2 larvae (IBSP 11303C), Sete Barras, state of São Paulo (24°23′16′′S, 47°55′33′′W), 20-IV-2013, Oligoryzomys nigripes; 2 larvae (IBSP 11304A), same locality, 20-IV-2013, Oligoryzomys sp.; 2 larvae (IBSP 11305), same locality, 10-VIII-2013, Euryoryzomys russatus; 1 larva (IBSP 11365), same data; 4 larvae (IBSP 11366), same locality, 10-VIII-2013, Monodelphis iheringi; 2 larvae (IBSP 11367), same locality, 15-VIII-2013, Oxymycterus sp.; 2 larvae (IBSP 11369), same locality, no date, Delomys sp.; 1 skin of larva and 3 deutonymphs (IBSP 12138B), Morro Grande, Cotia, state of São Paulo, no date, Euryoryzomys russatus; 1 skin of larva and 3 deutonymphs (IBSP 12140), same locality, no date, Akodon sp.; 1 deutonymph (IBSP 12141), same locality, no date and host information; 1 skin of larva (IBSP 12144), same locality, 15-IV-2015, Akodon sp.; 3 skins of larva and 3 deutonymphs (IBSP 12146A), same locality and date, Monodelphis sp.; 2 skins of larva and 2 deutonymphs (IBSP 12554A), same locality, no date and host information; 27 larvae, 15 skins of larva and 11 deutonymphs (IBSP 12557B), same locality, 20-VIII-2015, Akodon montensis; 4 skins of larva (IBSP 12559), same locality, no date and host information; 12 skins of larva (IBSP 12560), same locality, no date and host information; 1 skin of larva (IBSP 12695), 23-X-2013, Thaptomys nigrita.

Although expected size amplicons were obtained for 14/24 of the samples submitted to PCR, only three larvae and two deutonymphs yielded bands strong enough for sequencing. After an alignment of 421-bp including these five sequences, two haplotypes differing in one nucleotide were observed: one belonging to the deutonymphs and two larvae (GenBank accession numbers: KY934462, KY934464, MF113412 and MF113413), and another to one larva (KY934463). By BLAST analyses, both haplotypes of Q. brasiliensis were 96 – 99% similar to the sequences available in GenBank for Allothrombium sp. (KP276493) and Eutrombicula splendens (Ewing, 1913) (KP325057) respectively. For the gene COI, none of the tested samples amplified.
According to the original description of this species by Goff and Gettinger (1989), the holotype and paratypes of Q. brasiliensis were deposited in the BPBM, MZUSP, OMNH and NMNH collections. However, a search of these collections revealed no type specimens. Nevertheless, in the NMNH we found additional slide-mounted material coinciding with the same data of collection for the type specimens, except for the collecting date. After a review of all of the additional material, we found variation only in the position of the dorsal, preanal and postanal setae, depending on the degree of engorgement of the mites, as well as in the total number of idiosomal setae.
We have succeeded in obtaining deutonymphs, by rearing larvae on activated charcoal as substrate in order to remove plaster of Paris toxicity (Shatrov 2003). A great variety of substrate types have been tested successfully by other researchers (Wharton 1946; Wharton and Carver 1946; Michener 1946; Jenkins 1947; Farrel and Wharton 1949; Hyland 1951, 1961; Lipovsky 1953, 1954; Kaufman and Traub 1966; Nadchatram 1968; Everett et al. 1973; Mallow and Crossley 1984; Takahashi et al. 1993; Shatrov 1996; Tuegel and Wrenn 1998), including modifications in the proportion of activated charcoal, density of those substrates and sizes or kind of the culture containers.
In the present study, the deutonymphs of Q. brasiliensis were obtained under temperatures of 26°C (± 1°C), as well as the substrate with activated charcoal (1:1) demonstrated to be more efficient than in other proportion. Thus, the following parameters proved to be determinants of successful acquisition of deutonymph stages of Q. brasiliensis in laboratory conditions. The relative humidity inside the dish must be near the saturation level without being completely saturated in order to avoid the formation of fungi colonies that would need to be removed immediately.
Lipovsky (1951) tested different types of food, such as eggs of Lepidoptera, Coleoptera, Collembola, and Diptera for Eutrombicula alfreddugesi (Oudemans, 1910) and Eutrombicula splendens (Ewing, 1913), noticing that the preferred food source varies depending of the trombiculid species. Specimens of T. alfreddugesi and T. splendens fed on eggs Collembola as well as of Diptera; whereas Hirsutiella zachvatkini (Schluger, 1948) (Shatrov 2003), Eutrombicula cinnabaris (Ewing, 1920) (Tuegel and Wrenn 1998), Neoschongastia americana (Hirst, 1921) (Everett et al. 1973), Leptotrombidium pallidum (Nagayo, Miyagawa, Mitamura and Tamiya, 1919) (Takahashi et al. 1993), and Hannemania hegeneri Hyland, 1956 fed almost exclusively on Sinella curviseta (Collembola) eggs, in laboratory rearing. Eggs of Onychiurus sp. (Collembola) were given daily (at least 10 eggs per day) presented pink-like color (by fungi infection), and this fact was previously recorded by Lipovsky (1951). This author verified that eggs were infected with Penicillium, caused progressive lethargy which led to death of the specimens of E. alfreddugesi and E. splendens.
Temperature control was not a high priority issue for colony maintenance of Eutrombicula batatas (Linnaeus, 1758) and Blankaartia alleei (Ewing, 1926), since temperature variations of ± 5°C (Jenkins, 1947) did not affect the success of the colony. Shatrov (2003) observed the same situation for H. zachvatkini.
The only record of Q. brasiliensis was the type locality described from the Brasília (Federal District), collected on rodents H. megacephalus (cited as Oryzomys capito) and on marsupials G. agilis (cited as Marmosa agilis) and M. americana (Goff and Gettinger 1989). In this study, the species was recorded in the southern region of Adrianópolis (state of Paraná), and E. russatus was a new host record. We also added the cities of São Paulo, Cotia, Itapevi, Peruíbe and Sete Barras as new occurrences (Figure 5), and rodents of the species: A. montensis, C. paca, O. nigripes and T. nigrita; and marsupials of the species D. aurita and M. iheringi as new hosts for Q. brasiliensis.
Partial sequences obtained for the 18S ribosomal RNA gene of two larvae and two deutonymphs were identical between each other, and one larva yielded a sequence differing only in one nucleotide. BLAST analyses sequences of Q. brasiliensis were highly similar (99%) to E. splendens, the comparison of the complete sequences of both species should be done in order to draw further conclusions. These sequences correspond to the first molecular identification of the genus Quadraseta.
Kampen (2004) succeeded in amplifying the COI gene of Neotrombicula autumnalis (Shaw, 1790) larvae using primers for Tetranychidae mites designed by Navajas (1994); moreover, Moniuszko et al. (2015) were successful in amplifying the same gene, from larvae and deutonymphs of Hirsutiella zachvatkini (Schluger, 1948), using the primers by Dabert et al. (2008; 2010). In the present study, both pairs of primers were tested and none of them amplified the COI gene for Q. brasilensis.
This is the first description and record of Q. brasiliensis deutonymphs that developed from larvae maintained in laboratory conditions. The only previous description of the deutonymph instar of a chigger species recorded for Brazil was that of Eutrombicula ralphaudyi Vercammen-Grandjean and André, 1966.
To Marcelo Duarte da Silva, curator for invertebrates of the MZUSP, for the information about the holotype. To Katrina Menard, staff curator, and Janet Braun, head curator of the OMNH, and Jim Boone curator of the BPMH for the information about the paratypes. To Lee Goff and Donald Gettinger for helping us to try to find the missing type specimens. To Barry M. OConnor for his help on information on the species. To Gabrielle Ribeiro de Andrade and Maria Cristina Ferreira do Rosário for their technical contribution (CNPq no. 377343/2015-3 and 377342/2015-7, respectively). To Debra D. Creel, SEL-USDA, Floyd Shockley and David Furth, NMNH for technical assistance with the chigger specimens and literature. To D. Zeppelini, who provided the methodology for the maintenance of the collembolan colony and the identification of species of Collembola. To the Smithsonian NMNH, the Systematic Entomology Laboratory (USDA-ARS) for their support and material used in this study. To Marcelo Bahia Labruna, Departamento de Medicina Veterinária Preventiva e Saúde Animal (FMVZ-USP) for the molecular laboratory infrastructure. This work was supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq no. 454907/2014-1 and 440639/2015-8 to DMB-B, 377976/2014-8 to FCJ and 142409/2015-4 JAM-R) and Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP no. 2010/51875-9 to DMB-B and 2017/01416-7 to RB-S). SML was funded by CONICYT Programa de Formación de Capital Humano Avanzado, Beca Chile Nº 72140079. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the USDA; USDA is an equal opportunity provider and employer.
Altschul S.F., Gish W., Miller W., Myers E.W., Lipman D.J. 1990. Basic local alignment search tool. J. Mol. Biol., 215: 403-410. doi:10.1016/S0022-2836(05)80360-2 ![]()
Audy J.R. 1954. Malaysian Parasites IX. Notes on the taxonomy of trombiculid mites with description of a new subgenus. Stud. Inst. Med. Res. Malaya., 26: 123-170.
Bolton S.J., Klompen H., Bauchan G.R., Ochoa R. 2014. A new genus and species of Nematalycidae (Acari: Endeostigmata). J. Nat. Hist., 48: 1359-1373. doi:10.1080/00222933.2013.859318 ![]()
Brennan J.M. 1970. Two new genera and a new species of Neotropical chiggers (Acarina: Trombiculidae). Ann. Entomol. Soc. Am., 63: 1694-1696. doi:10.1093/aesa/63.6.1694 ![]()
Brennan J.M., Goff M.L. 1977. Keys to the genera of chiggers of the western hemisphere (Acarina: Trombiculidae). J. Parasitol., 63: 554-566. doi:10.2307/3280021 ![]()
Brennan J.M., Jones E.K. 1960. Chiggers of Trinidad, BWI (Acarina: Trombiculidae). Acarologia, 2: 493-540.
Brennan J.M., Jones E.K. 1964. Four new chiggers from Argentina (Acarina: Trombiculidae). J. Parasitol., 50: 698-702. doi:10.2307/3276133 ![]()
Crossley D.A. 1960. Comparative external morphology and taxonomy of nymphs of the Trombiculidae (Acarina). Univ. Kans. sci. bull., 15: 135-321.
Chomkzynski P.A. 1993. Reagent for the single-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. Biotechniques, 15: 532-537.
Dabert J., Ehrnsberger R., Dabert M. 2008. Glaucalges tytonis sp. n. (Analgoidea, Xolalgidae) from the barn owl Tyto alba (Strigiformes, Tytonidae): compiling morphology with DNA barcode data for taxon descriptions in mites (Acari). Zootaxa, 1719: 41-52.
Dabert M., Witalinski W., Kazmierski A., Olszanowski Z., Dabert J. 2010. Molecular phylogeny of acariform mites (Acari, Arachnida): Strong conflict between phylogenetic signal and long-branch attraction artifacts. Mol. Phylogenet. Evol., 56: 222-241. doi:10.1016/j.ympev.2009.12.020 ![]()
Dowling A.P.G., Bauchan G.R., Ochoa R., Beard J.J. 2010. Scanning electron microscopy vouchers and genomic data from an individual specimen: Maximizing the utility of delicate and rare specimens. Acarologia, 50: 479-485. doi:10.1051/acarologia/20101983 ![]()
Everett R.E., Price M.A., Kunz S.E. 1973. Biology of the chigger Neoschöngastia americana (Acarina: Trombiculidae). Ann. Entomol. Soc. Am., 66: 429-435. doi:10.1093/aesa/66.2.429 ![]()
Ewing H.E. 1913. New acarina. Part I. General considerations and descriptions of new species from Minnesota, Wisconsin, and Michigan. Bull. Am. Mus. Nat. Hist., 32: 93-122.
Ewing H.E. 1920. The genus Trombicula in America and the Orient. Ann. Entomol. Soc. Am., 13: 381-390. doi:10.1093/aesa/13.4.381 ![]()
Ewing H.E. 1926. Key to known adult Trombiculas (Adults of Chiggers) of the New World with descriptions of two new species (Acarina, Trombidoidea). Entomol. News, 37: 111-113.
Farrell C.E., Wharton G.W. 1949. A culture medium for chiggers (Acari: Trombiculidae). J. Parasitol., 35: 435. doi:10.2307/3273438 ![]()
Goff M.L., Gettinger D. 1989. Two new species of Schoengastiine chiggers (Acari: Trombiculidae) from Brazil and rediagnosis of Arisocerus Brennan, 1970. J. Med. Entomol., 26: 554-558. doi:10.1093/jmedent/26.6.554 ![]()
Goff M.L., Loomis R.B., Welbourn W.C., Wrenn W.J. 1982. A glossary of chigger terminology (Acari: Trombiculidae). J. Med. Entomol., 19: 221-238. doi:10.1093/jmedent/19.3.221 ![]()
Hirst S. 1921. On three new parasitic mites (Leptus, Schoengastia, and Demodex). Ann. Mag. Nat. Hist., 17: 37-39. doi:10.1080/00222932108632487 ![]()
Hyland R.E. 1951. Observations on the chigger mite Trombicula (Eutrombicula) splendens Ewing (Acarina: Trombiculidae). Ann. Entomol. Soc. Am., 44: 297-301. doi:10.1093/aesa/44.3.297 ![]()
Hyland R.E. 1956. A new species of chigger mite, Hannemania hegeneri Acarina: Trombiculidae). J. Parasitol., 42: 176-79. doi:10.2307/3274733 ![]()
Hyland R.E. 1961. Parasitic phase of the chigger mite Hannemania hegeneri on experimentally infested amphibians. Exp. Parasitol., 11: 212-25. doi:10.1016/0014-4894(61)90027-3 ![]()
International Commission on Zoological Nomenclature 1999. International code of zoological nomenclature, fourth edition. London: Int. Trust Zool. Nomencl. pp. 306.
Jenkins D.W. 1947. A laboratory method of rearing chiggers affecting man (Acarina: Trombiculidae). Ann. Entomol. Soc. Am., 40: 56-68. doi:10.1093/aesa/40.1.56 ![]()
Kampen H., Schöler A., Metzen M., Oehme R., Hartelt K., Kimmig P., Maier W.A. 2004. Neotrombicula autumnalis (Acari, Trombiculidae) as a vector for Borrelia burgdorferi sensu lato?. Exp. Appl. Acarol., 33: 93-102. doi:10.1023/B:APPA.0000029975.92510.90 ![]()
Kearse M., Moir R., Wilson A., Stones-Havas S., Cheung M., Sturrock S., Buxton S., Cooper A., Markowitz S., Duran C., Thierer T., Ashton B., Meintjes P., Drummond A. 2012. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics, 28: 1647-1649. doi:10.1093/bioinformatics/bts199 ![]()
Linnaeus C. 1758. Systema Naturae. Stockholm: Laurentius Salvius. pp. 824.
Lipovsky L.J. 1951. Collembola as food for chiggers. J. Parasitol., 37(3): 324-326. doi:10.2307/3273223 ![]()
Lipovsky L.J. 1953. Improved technique for rearing chigger mites (Acarina: Trombiculidae). Entomol. News, 64: 4-7.
Lipovsky L.J. 1954. Studies of the food habits of postlarval stages of chiggers (Acarina, Trombiculidae). Univ. Kansas Sci. Bull., 36: 943-958.
Kaufmann T., Traub R. 1966. Preliminary notes on parthenogenesis in a species of Trombiculid mites, Leptotrombidium arenicola Traub, 1960 (Acarina; Trombiculidae). J. Med. Entomol., 3(3-4): 256-258. doi:10.1093/jmedent/3.3-4.256 ![]()
Kethley J.B. 1990. Acarina: Prostigmata (Actinedida). In: Dindal, D. L. (Eds). Soil Biology Guide. John Wiley & Sons, New York, p. 667-756.
Mallow D., Crossley Jr D.A. 1984. Evaluation of five techniques for recovering postlarval stages of chiggers (Acarina: Trombiculidae) from soil habitats. J. Econ. Entomol., 77: 281-84. doi:10.1093/jee/77.1.281 ![]()
Michener C.D. 1946. Observations on the habits and life history of a chigger mite, Eutrombicula batatas (Acarina: Trombiculinae). Ann. Entomol. Soc. Am., 39(1): 101-118. doi:10.1093/aesa/39.1.101 ![]()
Miyajima M., Okumura T. 1917. On the life cycle of the akamushi carrier of Nippon river fever. Kitasato Arch. Exp. Med., 1: 1-14.
Moniuszko H., Zaleśny G., Mąkol J. 2015. Host-associated differences in morphometric traits of parasitic larvae Hirsutiella zachvatkini (Actinotrichida: Trombiculidae). Exp. Appl. Acarol., 67: 123-133. doi:10.1007/s10493-015-9925-0 ![]()
Nadchatram M. 1968. A technique for rearing trombiculid mites (Acarina) developed in a tropical laboratory. J. Med. Entomol., 5(4): 465-469. doi:10.1093/jmedent/5.4.465 ![]()
Nagayo M., Miyagawa Y., Mitamura T., Tamiya T. 1919. Über die sagen, zwei Formen der Tsutsugamushi nebst einer dritten, neuen. Form. Trans. Jpn. Path. Soc., 9: 107-110.
Navajas M., Gutierrez J., Bonato O., Bolland H.R., Mapangou-Divassa S. 1994. Intraspecific diversity of the cassava green mite Mononychellus progressivus (Acari: Tetranychidae) using comparisons of mitochondrial and nuclear ribosomal DNA sequences and cross breeding. Exp. Appl. Acarol., 18: 351-360. doi:10.1007/BF00116316 ![]()
Otto J.C., Wilson K.J. 2001. Assessment of the usefulness of ribosomal 18S and mitochondrial COI sequences in Prostigmata phylogeny. In: Halliday RB, Walter DE, Proctor HC, Norton RA, Colloff J (eds). Acarology: Proceedings of the 10th International Congress; Melbourne: CSIRO Publishing. p. 100-109.
Oudemans A.C. 1910. Acarologische Aanteekeningen XXXIII. Entomol. Ber. (Amst.), 3: 83-90.
Schluger E.G. 1948. [New red mites (Acari, Trombidiidae) of the fauna of the USSR]. Entomologicheskoe Obozrenie, 30: 157-164 (in Russian).
Shatrov A.B. 1996. Some features of life cycle and biology in trombiculid mites (Acariformes, Trombiculidae) under laboratory conditions. Zool. Zhurnal., 75: 677-691.
Shatrov A.B. 2003. Further observations on the life cycle and life strategy of a trombiculid mite, Hirsutiella zachvatkini (Schluger, 1948) (Acariformes: Trombiculidae), in the Laboratory. Acarina, 11(1): 73-90.
Shaw G. 1790. Natural vivarii. Nat' s Misc., 2: 42.
Stekolnikov A.A. 2008. Two new species of chigger mites (Acari: Trombiculidae) close to Neotrombicula minuta, application of nonlinear multivariate statistics. Acarina, 16: 21-29.
Stekolnikov A.A., Daniel M. 2012. Chigger mites (Acari: Trombiculidae) of Turkey. Zootaxa, 3216: 1-104.
Takahashi M., Machida K., Murata M., Misumi H., Hori E., Kawamura A.Jr., Tanaka H. 1993. Seasonal development of Leptotrombidium pallidum (Acari: Trombiculidae) observed by experimental rearing in the natural environment. J. Med. Entomol., 30(2): 320-325. doi:10.1093/jmedent/30.2.320 ![]()
Thompson J.D., Higgins D.G., Gibson T.J. 1994. CLUSTAL W: improving the sensitivity of progressive multiple sequence align- ment through sequence weighting, positions-specific gap penalties and weight matrix choice. Nucl. Acids Res., 22: 4673-4680. doi:10.1093/nar/22.22.4673 ![]()
Tuegel M.A., Wrenn W.J. 1998. Sexual dimorphism in morphology and development of the pest chigger, Eutrombicula cinnabaris (Acari: Trombiculidae). Int. J. Acarol., 24: 199-211. doi:10.1080/01647959808683585 ![]()
Vercammen-Grandjean P.H., Andre M. 1966. Nouveau Trombicula Adulte Originaire du Brésil Eutrombicula (Eutrombicula) ralphaudyi n. sp.. Acarologia, 8(1): 49-54.
Walter D.E., Krantz G.W. 2009. Chapter 7: Collecting, rearing, and preparing specimens. In: Krantz G.W.,Walter D.E. (eds.). A manual of acarology. Lubbock, Texas: Tech University Press. p. 83-96.
Wharton G.W. 1946. Observations on Ascoschöngastia indica (Hirst 1915) (Acarinida: Trombiculidae). Ecol. Monogr., 16(3): 151-184. doi:10.2307/1948648 ![]()
Wharton G.W., Carver R.K. 1946. Food of nymphs and adults of Neoschöngastia indica (Hirst, 1915). Science, 104: 76-77. doi:10.1126/science.104.2691.76 ![]()
Wohltmann A., Kohler J., Martin P. 2006. Endoparasitic mite infections of anuran amphibians from Bolivian montane rain forests, with descriptions of two new Hannemania species (Acari: Parasitengona: Trombiculidae: Leeuwenhoekiinae). Org. Divers. Evol., 6: 141-150. doi:10.1016/j.ode.2005.07.003 ![]()
Wohltmann A., Preez L., Rodel M.O., Kohler J., Vences M. 2007. Endoparasitic mites of the genus Endotrombicula Ewing, 1931 (Acari: Prostigmata: Parasitengona: Trombiculidae) from African and Madagascan anurans, with description of a new species. Folia Parasitol., 54: 225-235. doi:10.14411/fp.2007.031 ![]()
Yunker C.E., Brennan J.M. 1964. Four new chiggers (Acarina: Trombiculidae) from rodents of the epidemic area of Bolivian hemorrhagic fever. J. Med. Entomol., 1: 192-195. doi:10.1093/jmedent/1.2.192 ![]()



2017-06-02
Date accepted:
2017-11-23
Date published:
2018-03-22
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Jacinavicius, F. C.; Bassini-Silva, R.; Mendoza-Roldan, J. A.; Muñoz-Leal, S.; Hingst-Zaher, E.; Ochoa, R.; Bauchan, G. R. and Barros-Battesti, D. M.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)