



Description of two new species of Stigmaeopsis, Banks 1917 (Acari, Tetranychidae) inhabiting Miscanthus grasses (Poaceae)
Saito, Y.1 ; Sato, Y.2 ; Chittenden, A.R.3 ; Lin, J.-Z.4 and Zhang, Y.-X.5
1✉ Research Faculty of Agriculture, Hokkaido University, Sapporo 060-8589, Japan. Research Center of Engineer and Technology of Natural Enemy Resource of Crop Pest in Fujian, Institute of Plant Protection, Fujian Academy of Agricultural Sciences, Fuzhou 350013, China.
2Sugadaira Research Station, Mountain Science Center, University of Tsukuba, Ueda, Nagano 386-2204, Japan.
3Research Faculty of Fisheries Sciences, Hokkaido University, Hakodate, 041-8611, Japan.
4Research Center of Engineer and Technology of Natural Enemy Resource of Crop Pest in Fujian, Institute of Plant Protection, Fujian Academy of Agricultural Sciences, Fuzhou 350013, China.
5Research Center of Engineer and Technology of Natural Enemy Resource of Crop Pest in Fujian, Institute of Plant Protection, Fujian Academy of Agricultural Sciences, Fuzhou 350013, China.
2018 - Volume: 58 Issue: 2 pages: 414-429
https://doi.org/10.24349/acarologia/20184250ZooBank LSID: B2740172-4F3F-4FB0-8682-FB3E2E92A9B3
Keywords
Abstract
Stigmaeopsis celarius was first described by Banks (1917) from an introduced bamboo plant (Pleioblastus simonii) and established as the type species of the new genus Stigmaeopsis. Thirty-three years after the first description, this species was redescribed and Stigmaeopsis was synonymized with Schizotetranychus (Trägårdh 1915) by McGregor (1950). Much later, Saito et al. (2004) reinstated the genus Stigmaeopsis Banks (1917), described two new species and moved five others to the genus, formerly of the celarius species group of Schizotetranychus. Flechtmann (2012) added two new combinations, i.e. Stigmaeopsis malkovskii (Wainstein 1956) and Stigmaeopsis meghalayensis (Gupta and Gupta 1994). Furthermore, Saito et al. (2016) described two more Stigmaeopsis species from bamboo plants. Thus we now recognize 11 species as belonging to this genus.
Almost all members of the Stigmaeopsis genus have characteristic nest-weaving habits (Saito 2010; Saito et al. 2016). Considerable nest size variation exists between species on bamboo plants, which is correlated with species-specific variation in dorsal setal lengths (Mori and Saito 2013; Saito et al. 2016).
On the other hand, two Stigmaeopsis species are known to inhabit the perennial grasses, Miscanthus spp. and a reedgrass (Poaceae). In Stigmaeopsis miscanthi (Saito 1990), three forms, each of which possess different levels of male-to-male aggression (Saito 1995), were recently recognized (Sato et al. 2013a). The HG (males having high aggressiveness) and LW (males having low aggressiveness) forms of S. miscanthi have both been observed in Japan. The former is distributed over high temperature areas, and the latter over most low temperature areas with the exception of Hokkaido prefecture (Saito and Sahara 1999). However, there are very few morphological characters that can be used to identify these two forms other than the proportion of male leg development (armoured leg I in Saito and Sahara 1999; Sato et al. 2013a). Recently Sakamoto et al. (2017) found large differences between the LW, HG and a chinese (Cn) forms, suggesting that they are actually different species.
In the present study, we investigated the characteristics three forms in detail then discriminate two new species from S. miscanthi. Furthermore a new diagnosis of Stigmaeopsis was presented and a key to the world known species of Stigmaeopsis is also provided.
The two mite species described in the present study were collected from their respective fields (see results) and preserved in glass tubes containing MA solution (Saito and Osakabe 1993). Several of these preserved specimens were then prepared into permanent specimens using the Canada balsam method described in Saito et al. (1993). The specimens used for dorsal setae measurements were taken from colonies reared in the laboratory after collection and mounted on slides using Hoyer's solution. A 10g weight was gently placed on each cover glass to flatten the mite bodies. The specimens were kept on a hot plate at 55 °C for more than 7 days. The measurements were made from digital photographs taken with a digital camera (Sony Nex-7, Sony Corp., Tokyo, Japan) set up on a phase contrast microscope (Olympus BX63, Olympus Corp., Tokyo, Japan with 20x and 40x objective lens). Image J64 (provided by NIH, USA) for Macintosh (Apple Inc., USA) was used to measure seta and body lengths.
Diagnosis — Palp tarsus with six phaneres (projections): conical spinneret, two eupathidia, one solenidion and two tactile setae (n.b. seta b absent). Propodosoma well demarcated from hysterosoma; opisthonotum with 12 pairs of setae (c1-3, d1-2, e1-2, f1-2, h1-3; n.b. setae h1 are present, h3 placed ventrally); setae c1 widely spaced, in sublateral position close to setae c2 and c3; dorsal body setae slender with fine splits, length variable; opisthonotum with distinctive region of dorsocentral longitudinal striae between setae c1-c1 and d1-d1; two pairs of genital setae (g1-2); two pairs of pseudanal setae (ps1-2); tarsus I with two pairs of distal, adjacent duplex setae; tarsus II with one pair of duplex setae; empodium split into two claws, male tarsus I with empodium same as female empodium; male aedeagus weakly sigmoid, bending dorsally, without head. Leg setation typically reduced (e.g., genua 5-4-3-2).
Remarks — Stigmaeopsis is similar to some species of Schizotetranychus but is distinguished by its distinctive region of mid-dorsal longitudinal striae (though S. meghalayensis shows transverse striae), the loss of the palpal tactile seta b, and conical shape of spinneret.
(Japanese name: Tomo-sugomori-hadani)
ZOOBANK: AB01B8D1-9CE9-473B-97A3-F06E920F936B ![]()
Stigmaeopsis miscanthi low aggresiveness form in Saito (1995); Saito (2000); Saito (2010); Saito & Sahara (1999); Saito et al. (1999); Saito et al. (2000); Saito et al. (2002); Saito et al. (2005); Saito et al. (2013b); Sakagami et al. (2009); Sakamoto et al. (2017); Sato et al. (2000a, b); Sato et al. (2008); Sato et al. (2013a,b); Yano et al. (2011)
Description
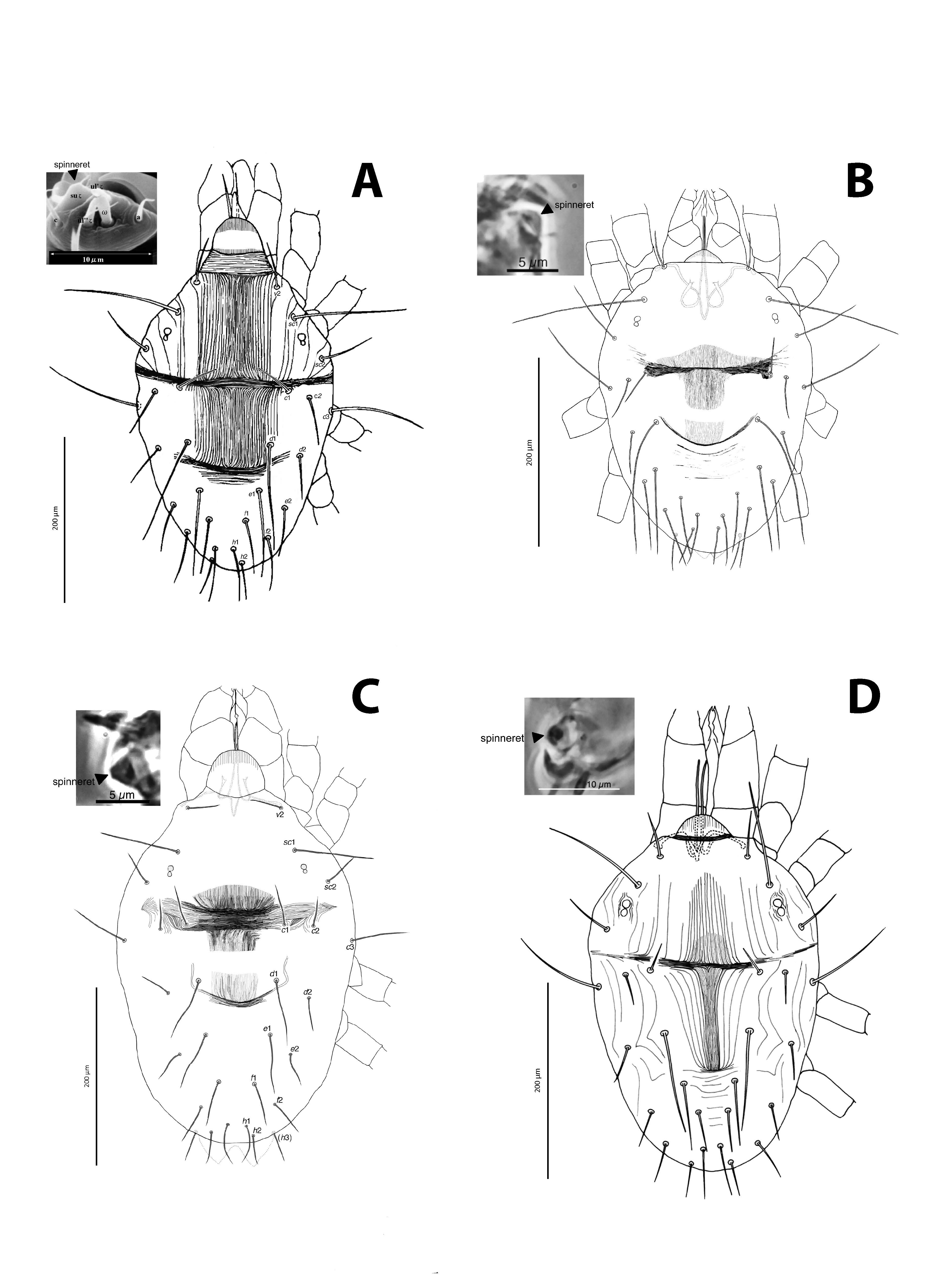
Female — Body flattened and wide, straw to greenish yellow with small blackish green spots. Body size 485.4 ± 15.9 μm from tip of rostrum to end of hysterosoma and 347.1 ± 15.7 μm from middle of horizontal line connecting setae v2 bases to end of hysterosoma. Peritreme bent and slightly dilated at distal end (but varies with specimen conditions). Propodosoma well demarcated from hysterosoma. Dorsal propodosomal setae sc1, 10% shorter than distance between their bases (Fig. 1, Table 1), and shorter than distance between bases of sc1 and c3. Bases of all dorsocentral hysterosomal setae (c1, d1, e1, f1) placed approximately in a straight line and the pair of lines forms a V shape. Length of dorsocentral hysterosomal setae d1 subequal with distance between their bases, just reaching bases of f1. Distance between bases of dorsolateral hysterosomal setae c2 longer than distance between bases of dorsolateral hysterosomal setae d2. Hysterosomal setae h3 exist ventrally. Lengths of dorsal setae and distances between their bases listed in Tables 1 and 2. Genital flap and area anterior to flap transversely striate (Fig. 1E). Distal segment of palpus has two simple setae and one spinneret (terminal sensilum), two eupathidia and one solenidion. Spinneret is conical in shape (Fig. 1B). Numbers of setae on leg segments presented in Figure 2 and Table 3.




Male — Body size 379.6 ± 39.6 μm from tip of rostrum to end of hysterosoma and 264.0 ± 26.7 μm from middle of horizontal line connecting setae v2 bases to end of hysterosoma. Lengths of dorsal setae and distances between their bases as in Tables 1 and 2. Femur-I with 9 tactile setae, one of which is tiny (dwarfed) and additional to those present in the female. Distal segment of palpus similar to female, spinneret slightly smaller (Fig. 1C). Aedeagus curved dorsally, weakly sigmoid (Fig. 1D). Numbers of setae on leg segments presented in Figure 3 and Table 3.








Type Material — HOLOTYPE: 1 male mounted with Canada balsam, Itoshima, Fukuoka, Japan, Miscanthus sinensis (Poaceae) on 12 May 2007, Saito Y. PARATYPE: 2 females and 2 males, same data; 2 females mounted with Canada balsam, Hiranai, Aomori, Japan, M. sinensis on 24 Aug. 2016 and 4 females and 1 male mounted with Hoyer's medium, Itoshima, Fukuoka, Japan, M. sinensis on 12 May 2007. These are deposited in the Hokkaido University Museum, Sapporo, 060-0810, Japan
Host and Distribution — On Miscanthus sinensis and Miscanthus spp. in Japan other than Hokkaido and Okinawa Prefectures.
Remarks — This species resembles Stigmaeopsis miscanthi but is distinguished from it by two morphological characters, as follows: the length of dorsal propodosomal setae sc1 shorter than distance between sc1 and c3 bases in S. sabelisi, but longer than that in S. miscanthi; The length of dorsocentral hysterodosomal setae d1 almost the same as distance between their bases in S. sabelisi, but much longer than that in S. miscanthi.
Life Type — This species lives within a dense web nest (WN-c life type in Saito 1983) built over depressions on the leaf adaxial surface (along midvein).
Etymology — Stigmaeopsis sabelisi is named in honor of the late Dr. Maurice W. Sabelis, a former Amsterdam University professor, who kindly advised on many of our long-term research projects.
(Japanese name: Bin-sugomori-hadani)
ZOOBANK: DCFE3743-86CD-45CC-BC50-9FB5F6DD8C32 ![]()
Stigmaeopsis miscanthi in Tsuji et al. (2011); Stigmaeopsis miscanthi Cn form in Sakamoto extslet al. (2017)




Female — Body flattened and wide, straw to greenish yellow with small blackish green spots. Body size 476.3 ± 25.2 μm from tip of rostrum to end of hysterosoma and 327.3.1 ± 15.0 μm from middle of horizontal line connecting setae v2 bases to end of hysterosoma. Propodosoma well demarcated from hysterosoma. Bases of all dorsocentral hysterosomal setae (c1, d1, e1 and f1) placed approximately in a straight line and the pair of lines forms a V shape. Length of dorsocentral hysterosomal setae d1 much longer than distance between their bases and exceed bases of f1. Distance between bases of dorsolateral hysterosomal setae c2 subequal to distance between bases of dorsolateral hysterosomal setae d2. Hysterosomal setae h3 (2nd para-anal setae) present. Lengths of dorsal setae and distances between their bases listed in Tables 1 and 2. Genital flap and area anterior to flap transversely striate (almost same as Fig. 1E). Palp tarsus with two simple setae and one conical spinneret , two eupathidia and one solenidion. Spinneret conical in shape (Fig. 4A). Numbers of setae on leg segments presented in Figures 5A-B and Table 3.
Male — Body size 415.9 ± 15.6 μm from tip of rostrum to end of hysterosoma and 302.4 ± 9.5 μm from middle of horizontal line connecting setae v2 bases to end of hysterosoma. Lengths of dorsal setae and distances between their bases as in Table 1. Femur-I with 9 normal setae, one of which is tiny (dwarfed) and additional to female setation. Numbers of setae and solenidia on distal segment of palpus presented in Fig. 4C and those on leg segments presented in Figures 5C-D, and Table 3. Aedeagus broadly curved dorsally, weakly sigmoid (Fig. 4D).
Type Material — HOLOTYPE: 1 male, made with Canada balsam, Fuqing, China, Miscanthus sp. 26 Feb. 2014, Saito Y. PARATYPES: 1 female, same data. 2 males (1 collected on 20 Feb. 2015 made with balsam and the other on 14 Apr. 2017 with Hoyer's medium), Fuzhou, China, Miscanthus sp. and 3 females (on 28 Aug. 2015, one with balsam and the other with Hoyer's medium), Fuzhou, China, Miscanthus sp. These are deposited in the Zoological Museum at Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, China.
Host and Distribution — Fuqing, Fuzhou and Wuiy, Fujian Prov., China parasitic to Miscanthus sinensis and Miscanthus spp.
Remarks — This species resembles both S. miscanthi and S. sabelisi but is distinguished from them by several characters as follows: the distance between the bases of dorsolateral hysterosomal setae c2 almost the same as that between the bases of dorsolateral hysterosomal setae d2 in S. continentalis, but the distance between the bases of c2 is longer than that between the bases of d2 in both S. sabelisi and S. miscanthi; in both S. continentalis and S. miscanthi, length of dorsocentral hysterosomal setae d1 are much longer than distance between their bases and extend past the bases of f1, but setae d1 are almost the same length as distance between their bases in S. sabelisi.
Life type — This species lives within dense web nests (WN-c life type in Saito 1983) built over depressions on leaf adaxial surface (along midvein).
space1cmDue to the addition of two new species, the key to the Stigmaeopsis species in Saito et al. (2016) should be changed as follows (following mite identification we recommend to refer to Figures 6 to 8 placed at the end of the article to get additional useful data on each species):
1. Length of setae d1 shorter than (or similar to) distance between bases of d1 setae members
...... 2
— Length of setae d1 longer than distance between bases of d1setae members
...... 8
2. Length of setae d1 shorter than (or similar to) distance between bases of d1 and e1 setae
...... 3
— Length of setae d1 longer than distance between bases of d1 and e1 setae
...... 5
3. Length of setae d2 longer than distance between bases of d1 and d2 setae
...... Stigmaeopsis tegmentalis Saito et Lin
— Length of seta d2 shorter than distance between bases of d1 and d2 setae
...... 4
4. Length of setae e1 similar to distance between bases of e1 and f1 setae
...... Stigmaeopsis saharai Saito et Mori
— Length of setae e1 much longer than distance between bases of e1 and f1 setae
...... Stigmaeopsis temporalis Saito et Ito
5. Length of seta e1 similar to distance between bases of e1 and e2 setae
...... Stigmaeopsis takahashii Saito et Mori
— Length of seta e1 longer than distance between bases of e1 and e2 setae
...... 6
6. Length of setae d1 similar to distance between bases of d1 and f1 setae
...... S. sabelisi* n. sp. Saito et Sato
— Length of setae d1 shorter than distance between bases of d1 and f1 setae
...... 7
7. Length of setae d1 shorter than distance between bases of d1 and e2 setae
...... Stigmaeopsis tenuinidus (Zhang et Zhang)
— Length of setae d1 longer than distance between bases of d1 and e2 setae
...... Stigmaeopsis malkovskii* (Wainstein)
8. Distance between bases of e1 setae similar to that between bases of d1 setae
...... Stigmaeopsis nanjingensis (Ma et Yuan)
— Distance between bases of e1 setae shorter than that between bases of d1 setae
...... 9
9. Length of setae d1 shorter than (or similar to) distance between bases of d1 and f2 setae
...... 10
— Length of setae d1 longer than distance between bases of d1 and f2 setae
...... 12
10. Distance between bases of c2 setae similar to that between bases of d2 setae
...... 11
— Distance between bases of c2 longer than that between bases of d2 setae
...... Stigmaeopsis miscanthi*(Saito)
11. Length of setae d2 shorter than (or similar to) distance between bases of d1 and d2 setae
...... Stigmaeopsis celarius Banks
— Length of setae d2 longer than distance between bases of d1 and d2 setae
...... S. continentalis* n. sp. Saito et Lin
12. Mediodorsal striae (central part of quadrilateral forming c1 and d1 setae bases) longitudinal
...... Stigmaeopsis longus (Saito)
— Mediodorsal striae (central part of quadrilateral forming c1 and d1 setae bases) transverse
...... Stigmaeopsis meghalayensis (Gupta et Gupta)
* mite species inhabiting Miscanthus sp. and reedgrasses.





Although S. meghalayensis does not satisfy several characters that represent Stigmaeopsis, we hold the decision until we can confirm the type specimen.The species marked with asterisk inhabit Miscanthus grasses and reedgrass (Poaceae), whereas all others inhabit Bambusoideae (Poaceae) plants. As shown in the above key, the lengths of dorsal setae are important characters for species identification in Stigmaeopsis. However, these setae are often broken and shortened in field collected mites, such that we must get our great attention to this possibility (Sakamoto et al. 2017). Specimens of newly emerged females from teleiocrysalis may help to avoid such troubles.
There are very few morphological differences between S. continentalis and S. miscanthi, thus they are considered to be cryptic species, such that geographic distribution (China in the case of the former and Japan in the case of the latter) and genetic information must be considered to distinguish S. continentalis from S. miscanthi. Furthermore, variation in the armored male morphology: S. miscanthi male leg I / leg III = 1.41; S. continentalis male = 1.32; S. sabelisi male = 1.29 (Saito and Sahara 1999; Sato et al. 2013a; in this study) may help with species identification, though males are rare in natural populations, due to their mortal combat tendencies.
From the above, we recognize 13 species of Stigmaeopsis. Although it is difficult to say that all species have been described, we reviewed several characteristics of Stigmaeopsis species in the genus diagnosis. From a morphological standpoint, no visible variations could be determined from the peritreme, aedeagus, striae of genital flap or those anterior to the flap (genital flap and area anterior to flap transversely striate) between congeneric species, which are useful characteristics for classifying other tetranychid species (Ehara 1999).
Almost all known species build nests over leaf depressions using silk threads (Saito et al. 2017; in this study) and live within them gregariously. Host plants are restricted to Poaceae distributed across Asian countries (India, Thailand, China (including Taiwan), Kazakhstan, Korea and Japan) other than artificial introduction.
We sincerely thank the late Maurice M. Sabelis who supported our work for long periods. We also thank Flechtmann C.H.W. who kindly provided us with several pieces of important literature. We also thank Gotoh T., Sahara K., Ito K., Uchida Y., Obokata S., Ji J., Chen X. and Sun L. for their help. This study was supported by the Fujian Council of Natural Science Foundation (2014J01108), China Recruitment Program of Global Experts (Foreign Experts) (2012-323), State Administration of Foreign Experts Affairs Key Project for Introduction of Foreign Expert (SZ2013003), the Agricultural Department of China (2017YFD0201000) and Special Research Fund of Fujian Academy of Agricultural Sciences (STIT2017-2-2). This study was also supported in part by JSPS KAKENHI Grant Numbers JP26891003 (YS), JP17K07556 (YS).
Banks N. 1917. New mites, mostly economic (Arach., Acar.). Entomol. News, 28: 193-199.
Ehara S. 1999. Revision of the spider mite family Tetranychidae of Japan (Acari, Prostigmata). Species Diversity, 4: 63-141.
Flechtmann C.H.W. 2012. Schizotetranychus-like spider mites (Acari, Prostigmata, Tetranychidae). revisited, new combinations and a key to groups of Schizotetranychus based on females. Acarologia, 52: 87-95. doi:10.1051/acarologia/20122039 ![]()
Gupta S.K., Gupta Y.N. 1994. A taxonomic review of Indian Tetranychidae (Acari: Prostigmata) with descriptions of new species, re-descriptions of known species and keys to genera and species. Genus 17. Schizotetranychus Trägårdh. Mem. Zool. Survey of India, 18: 86-99.
Ma E.-P., Yuan Y.-L. 1980. New species and new records of tetranychid mites from China I. (Acari: Tetranychidae). Acta Entmol. Sinica., 5: 42-45.
McGregor E.A. 1950. Mites of the family Tetranychidae. Am. Midl. Nat., 44: 257-420. doi:10.2307/2421963 ![]()
Mori K., Saito Y. 2013. Genetic basis of woven nest size in subsocial spider mites. Exp. Appl. Acarol., 60: 463-469. doi:10.1007/s10493-013-9661-2 ![]()
Saito Y. 1983. The concept of ``life types" in Tetranychinae. An attempt to classify the spinning behaviour of Tetranychinae. Acarologia, 24: 377-391.
Saito Y. 1990. Two new spider mite species of the Schizotetranychus celarius complex (Acari: Tetranychidae). Appl. Entomol. Zool., 25: 389-396. doi:10.1303/aez.25.389 ![]()
Saito Y. 1995. Clinal variation in male-to-male antagonism and weaponry in a subsocial mite. Evolution, 49: 413-417. doi:10.1111/j.1558-5646.1995.tb02273.x ![]()
Saito Y. 2010. Plant Mites and Sociality. Diversty and evolution. Tokyo, Springer, pp. 187. doi:10.1007/978-4-431-99456-5 ![]()
Saito Y., Ito K., Sakagami T. 2005. Imaginal induction of diapause in several `adult-female diapausing' spider mites. Physiol. Entomol., 30: 96-101. doi:10.1111/j.0307-6962.2005.00441.x ![]()
Saito Y., Kanazawa M., Sato Y. 2013. Life history differences between two forms of the social spider mite, Stigmaeopsis miscanthi. Exp. Appl. Acarol., 60: 313-320. doi:10.1007/s10493-012-9646-6 ![]()
Saito Y., Lin J.-Z., Zhang Y.-X., Ito K., Liu Q., Chittenden A.R. 2016. Two new species and four new life types in Tetranychidae. Ann. Entomol. Soc. Amer., 109: 463-472. doi:10.1093/aesa/sav158 ![]()
Saito Y., Mori K., Chittenden A.R. 1999. Body characters reflecting the body size of spider mites in flattened specimens (Acari, Tetranychidae). Appl. Ent. Zool., 34: 383-386. doi:10.1303/aez.34.383 ![]()
Saito Y., Mori K., Chittenden A.R., Sato Y. 2000. Correspondence of male-to-male aggression to spatial distribution of individuals in field populations of a subsocial spider mite. J. Ethol., 18: 79-83. doi:10.1007/s101640070004 ![]()
Saito Y., Mori K., Sakagami T., Lin J.-Z. 2004. Reinstatement of the genus Stigmaeopsis Banks, with descriptions of two new species (Acari, Tetranychidae). Ann. Entomol. Soc. Amer., 97: 635-646. doi:10.1603/0013-8746(2004)097[0635:ROTGSB]2.0.CO;2 ![]()
Saito Y., Sahara K. 1999. Two clinal trends in male-male aggressiveness in a subsocial spider mite. Behav. Ecol. Sociobiol., 46: 25-29. doi:10.1007/s002650050588 ![]()
Saito Y., Sakagami T., Sahara K. 2002. Differences in diapause attributes between two clinal forms distinguished by male-to-male aggression in a subsocial spider mite, Schizotetranychus miscanthi Saito. Ecol. Res., 17: 645-653. doi:10.1046/j.1440-1703.2002.00522.x ![]()
Saito Y., Osakabe Mh., Sakagami Y., Yasui Y. 1993. A method for preparing permanent specimens of mites with Canada balsam. Appl. Entomol. Zool., 28: 593-597. doi:10.1303/aez.28.593 ![]()
Saito Y., Zhang Y.-X., Mori K., Ito K., Sato Y., Chittenden A.R., Lin J.-Z., Chae Y., Sakagami T., Sahara K. 2016. Variation in nesting behavior of eight species of spider mites, Stigmaeopsis having sociality. Sc. Nat., 103: 87. doi:10.1007/s00114-016-1408-697 ![]()
Sakagami T., Saito Y., Kongchuensin M., Sahara K. 2009. Molecular phylogeny of Stigmaeopsis, with special reference to speciation through host plant shift. Ann. Entomol. Soc. Amer., 102: 360-366. doi:10.1603/008.102.0303 ![]()
Sakamoto H., Matsuda T., Suzuki R., Saito Y., Lin J.-Z., Zhang Y.-X., Sato Y., Gotoh T. 2017. Molecular identification of seven species of the genus Stigmaeopsis (Acari: Tetranychidae) and preliminary attempts to establish their phylogenetic relationship. Syst. Appl. Acarol., 22: 91-101. doi:10.11158/saa.22.1.10 ![]()
Sato Y., Breeuwer J.A.J., Egas M., Sabelis M.W. 2015. Incomplete premating and postmating reproductive barriers between two parapatric populations of a social spider mite . Exp. Appl. Acarol., 65: 277-291. doi:10.1007/s10493-015-9878-3 ![]()
Sato Y., Egas M., Sabelis M.W., Mochizuki A. 2013a. Male-male aggression peaks at intermediate relatedness in a social spider mite. Ecology and Evolution, 3: 2661-2669. doi:10.1002/ece3.661 ![]()
Sato Y., Sabelis M.W., Mochizuki A. 2013b. Asymmetry in male lethal fight between parapatric forms of a social spider mite. Exp. Appl. Acarol., 60: 451-461. doi:10.1007/s10493-013-9668-8 ![]()
Sato Y., Saito Y., Chittenden A.R. 2008. The parapatric distribution and contact zone of two forms showing different male-to-male aggressiveness in a social spider mite, Stigmaeopsis miscanthi (Acari: Tetranychidae). Exp. Appl. Acarol., 44: 265-276. doi:10.1007/s10493-008-9147-9 ![]()
Sato Y., Saito Y., Mori K. 2000a. Reproductive isolation between populations showing different aggression in a subsocial spider mite, Schizotetranychus miscanthi Saito (Tetranychidae: Acari). Appl. Ent. Zool., 35: 605-610. doi:10.1303/aez.2000.605 ![]()
Sato Y., Saito Y., Mori K. 2000b. Patterns of reproductive isolation between two groups of Schizotetranychus miscanthi Saito (Tetranychidae: Acari) showing different male aggression traits. Appl. Ent. Zool., 35: 611-618. doi:10.1303/aez.2000.611 ![]()
Tsuji N., Chittenden A.R., Ogawa T., Takada T., Zhang Y.-X., Saito Y. 2011. The possibility of sustainable pest management by introducing bio-diversity: simulations of pest mite outbreak and regulation. Sustain Sci., 6: 97-107. doi:10.1007/s11625-010-0113-1 ![]()
Trägårdh I. 1915. Bidrag till Kännedomen om Spinnkvalstren (Tetranychus Duf.). Meddelande Nr. 109 fran Centralanstalten för försöksväsendet pa Jordbruksomradet, 109 (Entomol. Avd. 20): 1-60.
Wainstein B.A. 1956. Material on the fauna of tetranychid mites of Kazakhstan. Tr. Resp. St. Zashch. Rast. Kazifilial Vaskhnil, 3: 70-83.
Yano J., Saito Y., Chittenden A.R., Sato Y. 2011. Variation in counterattack success against a phytoseiid predator between two forms of the social spider mite Stigmaeopsis miscanthi. J. Ethol., 29:337-342 doi:10.1007/s10164-010-0265-6 ![]()
Zhang Z.-Q., Zhang Y.-X., Lin J.-Z. 2000. Mites of Schizotetranychus (Acari: Tetranychidae) from moso bamboo in Fujian, China. Syst. Appl. Acarol. Spec. Pub., 4: 19-35. doi:10.11158/sasp.4.1.5 ![]()



2017-07-31
Date accepted:
2017-11-22
Date published:
2018-03-15
Edited by:
Auger, Philippe

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Saito, Y.; Sato, Y.; Chittenden, A.R.; Lin, J.-Z. and Zhang, Y.-X.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)