



A checklist of the water mites of Central Asia with description of six new species (Acari, Hydrachnidia) from Kyrgyzstan
Pešić, Vladimir1 and Smit, Harry2
1✉ Department of Biology, University of Montenegro, Cetinjski put b.b., 81000 Podgorica, Montenegro.
2Naturalis Biodiversity Center, P.O. Box 9517, 2300 RA Leiden, The Netherlands.
2018 - Volume: 58 Issue: 1 pages: 165-185
https://doi.org/10.24349/acarologia/20184236ZooBank LSID: FA671766-4010-466B-9AF8-681227A2A661
Keywords
Abstract
The water mites of Central Asia have been insufficiently studied (Table 1). This region, extending from Afghanistan in the south to Russia in the north and from the Caspian Sea in the west to China in the east, consists today of five countries: Kazakhstan, Kyrgyzstan, Tadjikistan, Turkmenistan, and Uzbekistan. The water mites from Central Asia have been investigated by several researchers, notably Daday (1903), Thor (1909, 1930), Ruzskij (1929), and Tjutenkow (1956). Furthermore, in the twenties and thirties of the last century, Iwan Sokolow greatly contributed to knowledge of water mites of this region. He described Torrenticola turkestanica from Uzbekistan (Sokolow 1926); examined the diversity of Bukhara (Sokolow 1928); described Hygrobates kirgizicus from a tributary of the Chui river in the Ortotokoy valley (Tian Shian Mountains) at altitudes of 1700-1800 m (Sokolow 1935); and he described many new species and a new genus (Tadjikothyas) from Tadjikistan (Sokolow 1948). Then, water mites of the region were untouched until 1973 when Jankovskaja described phreatic and spring fauna of northern Tian Shian (Kyrgyzstan) and southern Kyzylkum (Uzbekistan).





In the 21st century, Central Asian water mite research began with the description of two new Sperchon Kramer, 1877 from streams near Bishkek, Kyrgyzstan (Tuzovskij 2003). Recently, Smit (2016) described a new genus (Kazakhithyas) from a rheohelocrene spring from the Tien Shan Mountains, Kazakhstan. Finally, based on morphological and molecular data we described a new Neumania species from a pond near Issyk Kul Lake in Kyrgyzstan and recorded Limnesia koenikei for the first time from this area (Pešić & Smit 2017).
The present study is based on material collected by the authors during a joint collecting trip from 1-15 July 2013 to Kyrgyzstan, a landlocked country in Central Asia, bordering Kazakhstan, Tajikistan, Uzbekistan, and China. In this paper, descriptions of six new species are given.
Water mites were collected by hand netting, sorted on the spot from the living material and preserved in Koenike-fluid. Specimens were dissected as described in Gerecke et al. (2007) and slide mounted in Hoyer’s medium. Morphological nomenclature follows Gerecke et al. (2016). Holotypes and paratypes of the new species are deposited in the Naturalis Biodiversity Center in Leiden (RMNH).
The composition of the material is given as: males/females/deutonymphs. All measurements are given in μm. The following abbreviations are used: Ac-1 = first acetabulum; asl. = above sea level; Cx-I = first coxae; dL = dorsal length; H = height; I-L-4-6 = fourth-sixth segments of first leg; L = length; lL =lateral length; mL = medial length; NP = National Park; P-1– P-5 = palp segment 1-5; S-1 = proximal large ventral seta at I-L-5; S-2 = distal large ventral seta at I-L-5; Vgl-1 = ventroglandularia 1; W = width.
Subfamily Wandesiinae Schwoerbel, 1961
Genus Wandesia Schechtel, 1912
ZOOBANK: B8E7A868-F8ED-45A1-827E-3A8ECBB9C723 ![]()

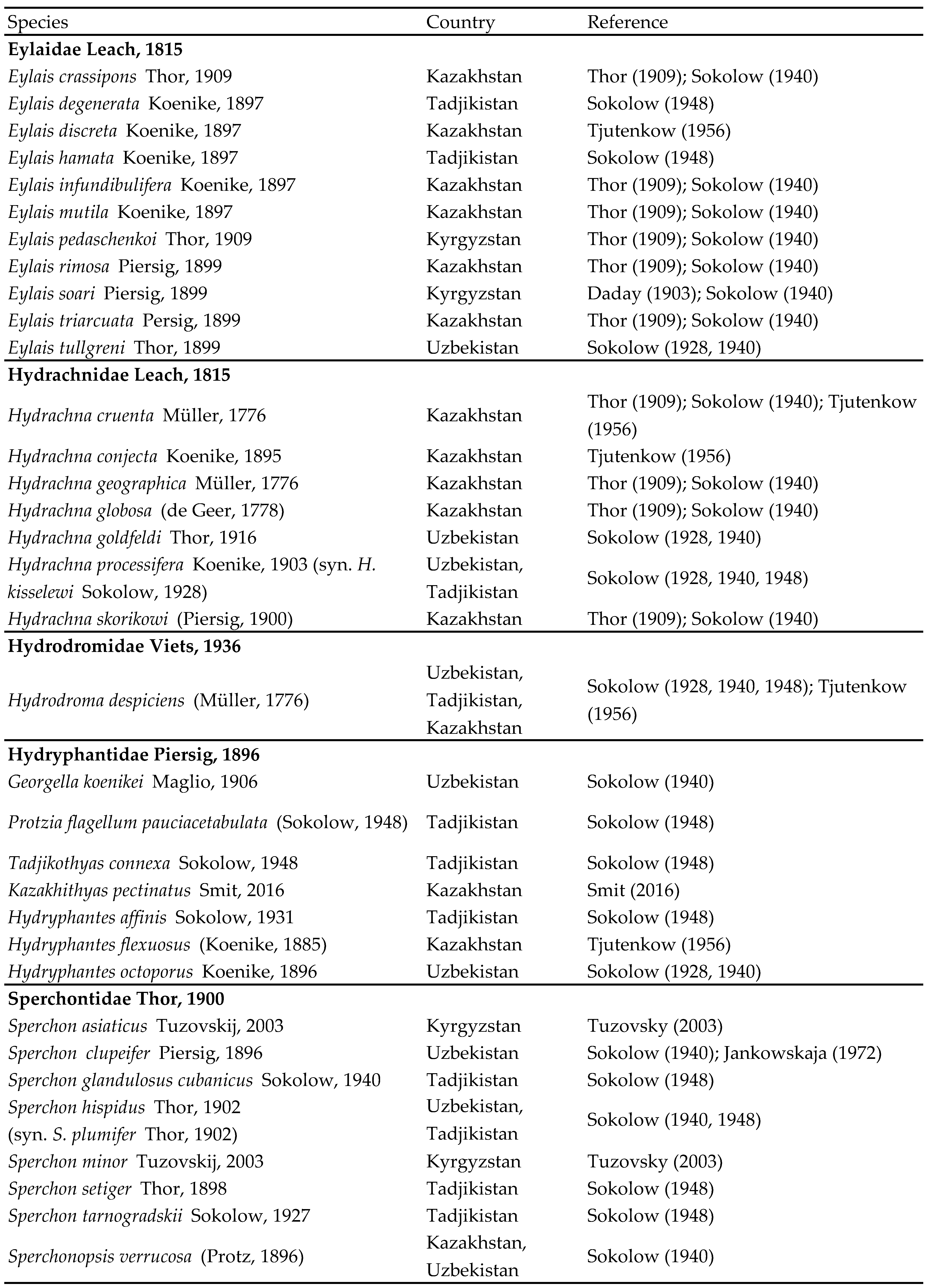
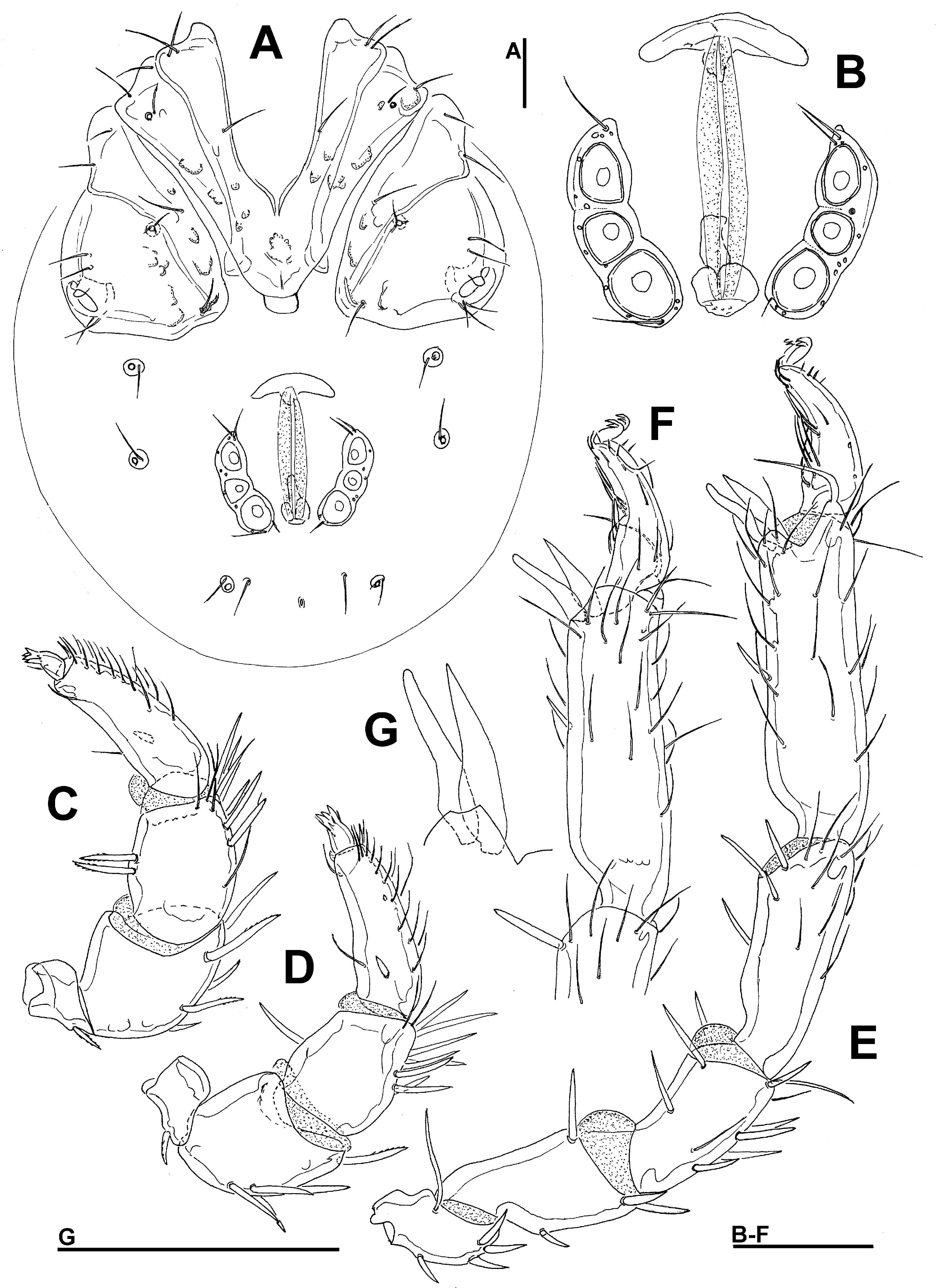
Type material — Holotype female (RMNH), Kyrgyzstan, KR15 Karakal region, road to May Saz Pass, fast flowing river, 42°25.004'N, 78°58.100'E, 2981 m asl., 12.vii.2013, leg. Pešić & Smit, dissected and slide mounted. Diagnosis — Female (Male unknown). Idiosoma small (L 1500 μm); Cx-IV reduced, with medial part not reaching medial part of Cx-III; P-4 slender, L/H ratio 3.7; P-5 without a flagellar seta; basal segment of chelicera curved; no visible pairs of setae inserting in the membranous integument around gonopore; Ac-1 much smaller than posterior pairs of acetabula; III-L-6 and IV-L-6 distally slightly enlarged.
Description — Female — Body elongated; lateral eyes unpigmented; integument reticulated. Cx-I+II with slightly convex medial margin, broadly rounded medioposteriorly, posterolaterally concave; Cx-III with broad medial edge; Cx-IV reduced (Figure 1A); Cx-I with three setae; Cx-II without setae; Cx-III and Cx-IV with one seta each. Genital field with three pairs of stalked elongated genital acetabula; distance between Ac-1 and Ac-2 three times longer than between Ac-2 and Ac-3 (Figure 1E); gonopore between first pairs of acetabula. Palp (Figures 1B-C): P-1 without setae; P-2 with one strong distal seta; P-3 with two slender dorsodistal setae; P-4 with large dorsodistal spine and one thin distal seta, ventral seta short, in distal third. Legs: III-L-6 and IV-L-6 distally slightly enlarged; claws simple hook-like.
Measurements — Idiosoma L 1494, W 363; Cx-I+II total L 116, W 78, Cx-III+IV lateral L 128; Ac-1 L 14, L/W ratio 1.13; Ac-2 L 27, L/W ratio 1.4, Ac-3 L 34, L/W ratio 1.59.
Gnathosoma — palp total L 230; dL/H, dL/H ratio: P-1, 19/30, 0.63; P-2, 50/34, 1.49; P-3, 49/36, 1.37; P-4, 89/24, 3.7; P-5, 23/12, 2.0; L ratio P-2/P-4 0.56. Gnathosomal base vL 125; chelicera total L 172, basal segment L 130, claw L 44, L basal segment/claw ratio 3.0.
Legs — dL of I-L: 42, 63, 80, 91, 102, 109. dL of II-L: 41, 63, 69, 88, 100, 114. dL of III-L: 43, 52, 63, 77, 91, 106. dL of IV-L: 38, 68, 81, 92, 100, 100.
Discussion — The new species is morphologically most similar to Wandesia reducta, a species described by Tuzovskij (1987) based on three females and a larva from Magadan region in the Far East of Russia and later on reported by Pešić et al. (2015) from South Korea. Recently Semenchenko (2016) described the male of this species. Both species share P-5 without a flagellar seta and Cx-IV reduced, with the medial part not reaching the medial part of Cx-III. The female of W. reducta (in parentheses data taken from Tuzovskij 1987) differs in larger idiosoma dimensions (L 2280-2610 μm), the genital field with 3-5 pairs of small setae located around the anterior half of gonopore, P-4 shorter (L 65 μm) and stouter, and III-L-6 and IV-L-6 distally strongly enlarged (see Tuzovskij 1987, figure 3I).
Etymology — In remembrance of our colleague Prof. Gerd Alberti (1943-2016).
Distribution — Kyrgyzstan; known only from the locus typicus (Figure 10E).
Genus Torrenticola Piersig, 1896
ZOOBANK: 9CE2E5A0-B7CA-453B-A6D5-8F17A2F6BD0B ![]()

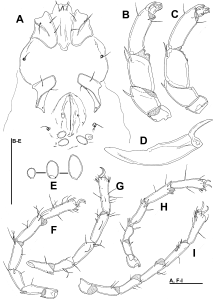
Type material — Holotype female (RMNH), Kyrgyzstan, KR8 Chon-Kemin NP, upper part of stream near Ashu resort, Kalman Ashu village, 42°42.276'N, 76°05.101'E, 1644 m asl., 10.vii.2013, leg. Pešić & Smit, dissected and slide mounted.
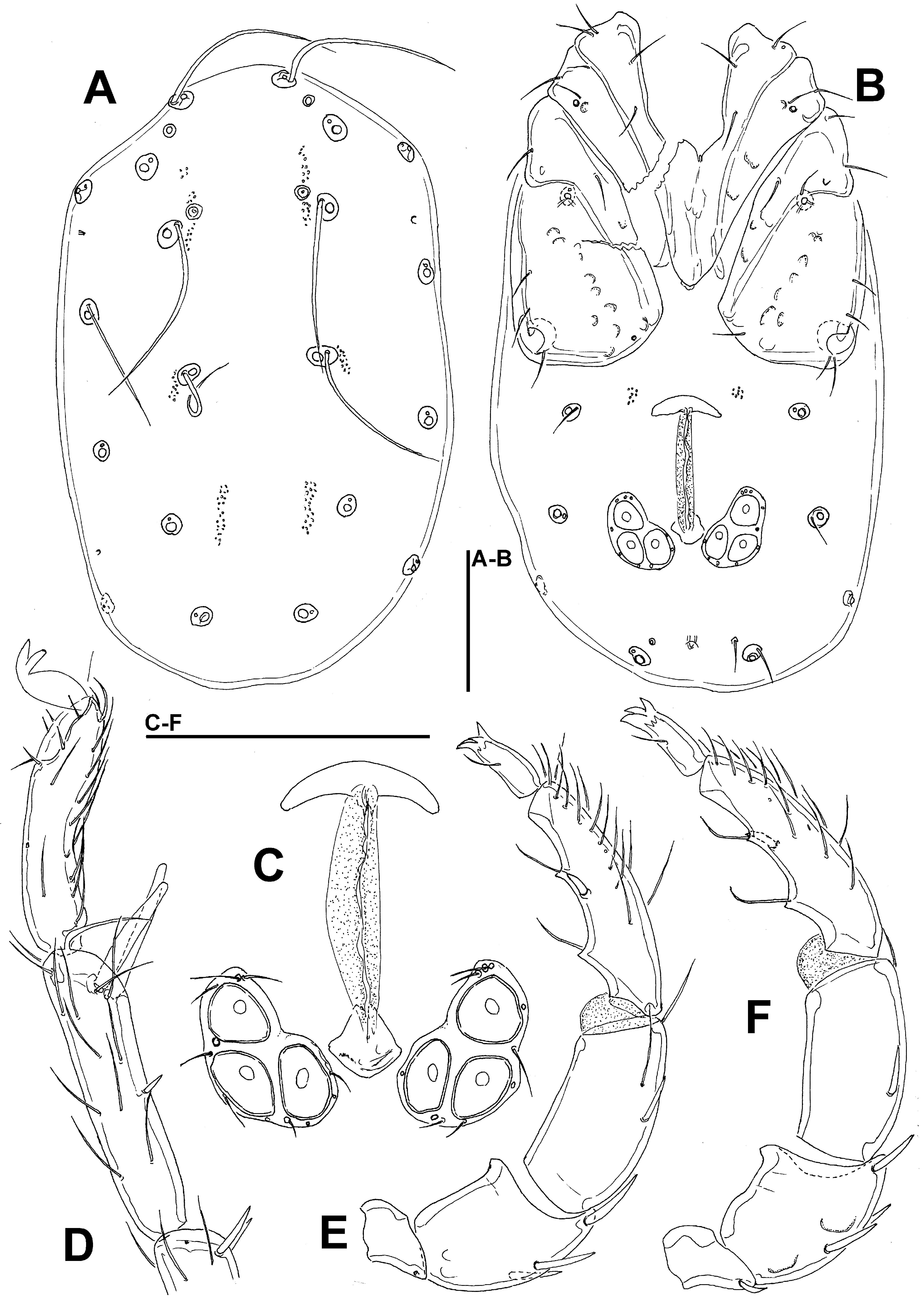
Diagnosis — Female (Male unknown). Idiosoma elongated (dorsal shield L/W ratio 1.6); Cxgl-4 subapical; medial suture line of Cx-II+III long (> 100 μm); P-2 with a laterally compressed, longish (> 30% of ventral margin), anteriorly directed ventrodistal extension; P-4 L/H ratio 4.6.
Description — Female — Idiosoma elongated; dorsal shield without colour pattern; gnathosomal bay U-shaped, proximally rounded; Cxgl-4 subapical, only slightly posterior to Cx-I tips; medial suture line of Cx-II+III relatively long; suture lines of Cx-IV extending posteriorly beyond posterior margin of genital field, laterally curved; genital field pentagonal in shape; excretory pore on the level with Vgl-2, both slightly away from the line of primary sclerotization; gnathosomal rostrum long (Figure 2E), ventral margin in lateral view curved; P-2 nearly equal in length as P-4, ventral margin of P-2 distally with a laterally compressed, longish (>30% of ventral margin), anteriorly directed and apically serrated hyaline extension and a very short seta laterally at base of projection; P-3 with a shorter, subrectangular, apically serrated ventrodistal projection, and a moderately long seta laterally at base of projection; P-4 slender, with ventral tubercles pointed and separated, bearing one long and three short setae (Figure 2D).
Measurements — Idiosoma (ventral view: Figure 2C) L 846, W 463; dorsal shield (Figure 2B) L 659, W 408, L/W ratio 1.6; dorsal plate L 619; frontal plate L 141-150, W 50, L/W ratio 2.8-3.0; shoulder plate L 191-192, W 66, L/W ratio 2.9; shoulder/frontal plate L 1.28-1.36. Gnathosomal bay L 150, Cx-I total L 294, Cx-I mL 142, Cx-II+III mL 116; ratio Cx-I L/Cx-II+III mL 2.54; Cx-I mL/Cx-II+III mL 1.23. Genital field L/W 169/149, ratio; distance genital field-excretory pore 184, genital field-caudal idiosoma margin 242.
Gnathosoma — palp total L 350; dL/H, dL/H ratio: P-1, 35/30, 1.17; P-2, 112/53, 2.1; P-3, 72/47, 1.53; P-4, 114/25, 4.56; P-5, 17/-, -; L ratio P-2/P-4 0.98. Gnathosomal base vL 378; chelicera total L 416.
Discussion — The combination of a flanged palp with subapical Cxgl-4, only slightly posterior of Cx-I tips, makes the new species similar to Torrenticola malaisei (Lundblad, 1941), T. prionipalpis (Lundblad, 1941), and T. lamellifera (Lundblad, 1941), all known from Burma (Lundblad 1969). The latter species has been synonymized by Wiles (1997) with T. malaisei, but recently this synonymization has been rejected by Pešić et al. (2012). Females of T. malaisei and T. lamellifera can be distinguished from those of T. kyrgyzica by their more roundish idiosoma, a short medial suture line of Cx-II+III, and a longer ventral seta on P-3 (see Wiles 1997). Moreover T. malaisei has a stouter P-4. Torrenticola prionipalpis (Lundblad, 1941), known from a single male, has a roundish idiosoma, a more slender P-4, a smaller ventral seta on P-2, and P-4 ventral tubercle, ending in two tips separated by a concavity, with four short setae (see Wiles 1997).
Etymology — Named after the country where the new species was found.
Distribution — Kyrgyzstan; known only from the locus typicus (Figure 10D).
Genus Atractides Koch, 1837
ZOOBANK: F96801F6-F88E-4206-87B4-4ABA0296BD75 ![]()
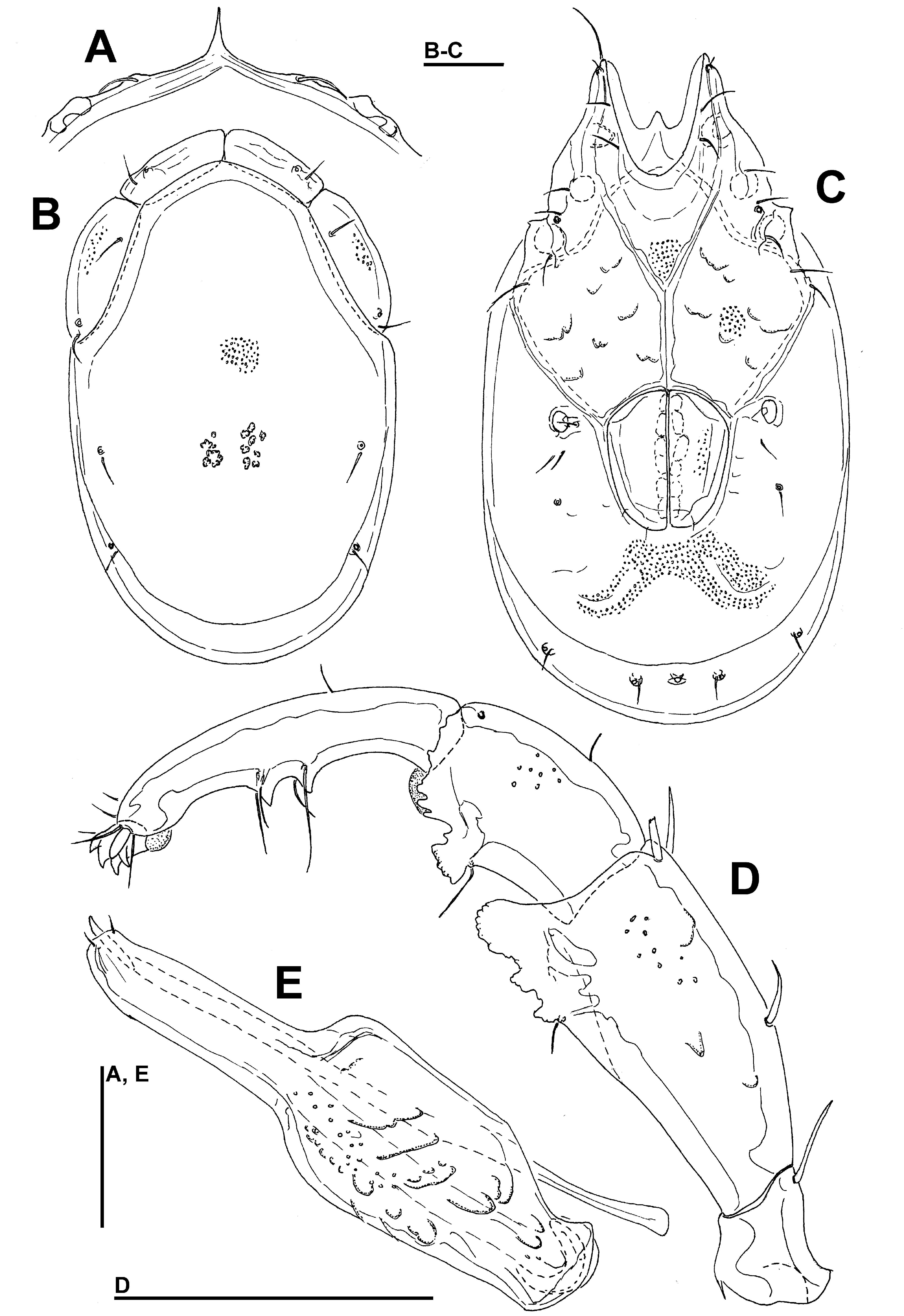
Type material — Holotype female (RMNH), Kyrgyzstan, KR10 Grigorievka Gorge, rheocrenic spring, 42°46.585'N, 77°28.391'E, 1960 m asl., 11.vii.2013, leg. Pešić & Smit, dissected and slide mounted. Paratype: one female, Kyrgyzstan, KR23 Karakal region, rheohelocrenic spring, Žeti-Egaz village, 42°19.238'N, 78°51.017'E, 2094 m asl., 13.vii.2013, leg. Pešić & Smit, left palp and I-L dissected and slide mounted.
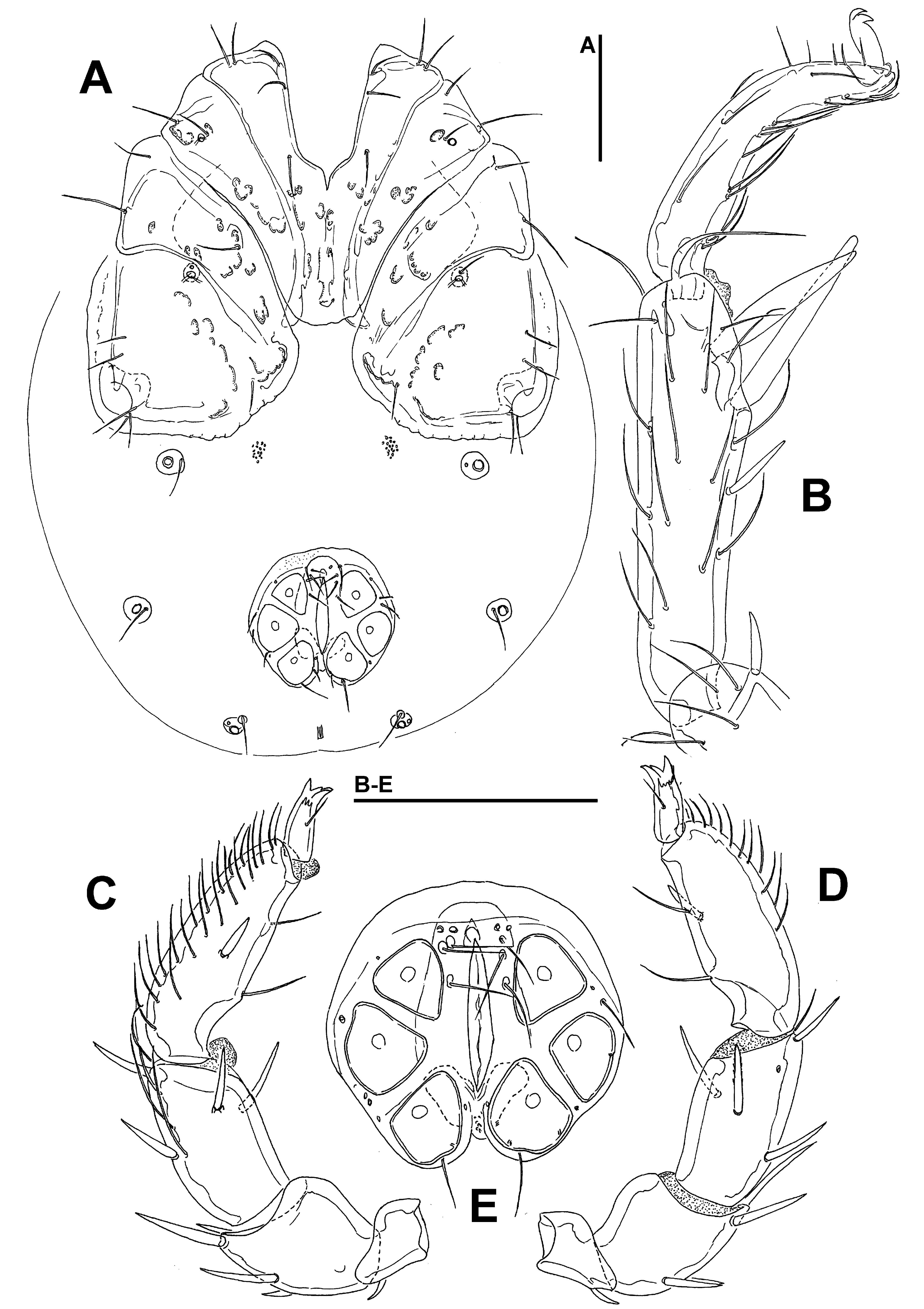
Diagnosis — Female (Male unknown). Integument striated; suture line Cx-III and –IV in its medial part directed posteriorly; P-3 ventral margin slightly convex with one (occasionally two) ventral seta; P-5 short; excretory pore smooth; acetabula in a slightly curved line; I-L-5 with S-1 ending in a bluntly pointed tip.
Description — Female — Integument dorsally striated, muscle insertions unsclerotized. Posterior margin of Cx-I weakly curved, with a subcuticular protrusion; suture line Cx-III and -IV in its medial part directed posteriorly (Figure 3A). Genital plates with narrow sclerite border, slightly indented between Ac-1/2 and Ac-2/3, acetabula in a slightly curved line (Figure 3B). Excretory pore smooth; Vgl-1 not fused to Vgl-2. P-3 ventral margin slightly convex, with one (occasionally two) ventral seta(e), inserted in the middle of segment; P-4 setae insertions dividing ventral margin into sectors 1:4:2, sword seta proximally from proximoventral seta; P-5 short (Figures 3C-D). I-L-5: S-1 with a blunt tip, more slender than S-2, S-2 pointed, strongly enlarged in the proximal part (Fig. 3G); I-L-6 stout, very thick at the base, distally tapering (Figures 3E-F).
Measurements — (holotype; in parentheses some measurements of paratype) — Idiosoma L 938 (1113), W 688 (925); coxal field: L 434; Cx-III W 553; Cx-I+II mL 134, lL 355; genital field L/W 223/224, genital plates L 144-145, pregenital sclerite W 122, maximum diameter Ac 1-3: 44, 31-38, 47-51.
Gnathosoma — palp total L 450; dL/H, dL/H ratio: P-1, 48/44, 1.1 (50/47, 1.06); P-2, 127/92, 1.38 (121/91, 1.34); P-3, 103/80, 1.29 (95/79, 1.2); P-4, 138/43, 3.2 (123/44, 2.82); P-5, 34/19, 1.83 (33/19, 1.75); L ratio P-2/P-4 0.92 (0.98). Chelicera total L 298.
Legs — I-L-5 dL 248 (232), vL 217 (186), dL/vL ratio 1.15 (1.25), maximum H 67 (72), dL/maximum H 3.7 (3.2), S-1 L 66, L/W ratio 5.3, S-2 L 70, L/W ratio 4.5, distance S-1-2 5.0, L ratio S-1/2 0.95; I-L-6 L 119 (108), maximum H 42 (41), L/maximum H ratio 2.8 (2.6); L ratio I-L-5/6, 2.1 (2.15).
Discussion — The new species is provisionally assigned to the subgenus Polymegapus K. Viets, 1926, which was considered by Gerecke (2003) to be a monophyletic clade. The new species does not fit the diagnosis of the subgenus given by Gerecke et al. (2016) in the following features: 1) suture line Cx-III and –IV in its medial part directed posteriorly, 2) Cx-4 without triangular laterocaudal extensions of secondary sclerite, 3) I-L-5 with ventral margin shorter than dorsal margin, 4) P-5 short and less slender, and 5) excretory pore unsclerotized. The combination of the characters given under 1), 3) and 5) make the new species close to A. putihi Wiles, 1991 from SE Asia (Wiles 1991, Pešić & Smit 2009, Smit & Pešić 2014). The latter species differs in genital plates, restricted to the level of the postgenital platelet, with the acetabula in a triangular arrangement, P-3 ventral margin slightly concave, without setae and IV-L-5 with 2 swimming setae. Probably A. grigorievka n. sp. represents an early derivative member with the typical palp and I-L not yet fully developed.
Etymology — Named after the canyon where the new species was found. The name is a noun in apposition.
Distribution — Kyrgyzstan, known from one rheocrenic (Figure 10C) and one rheohelocrenic spring (Figure 10G).
ZOOBANK: DA72B28F-BBD4-4EDC-B2B6-B4071B868FFB ![]()

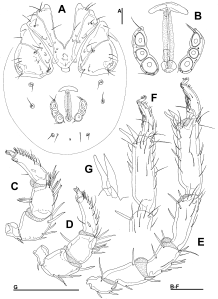
Type material — Holotype female (RMNH), Kyrgyzstan, KR22 Karakal region, stream, 42°44.342'N, 78°53.681'E, 1917 m asl., 13.vii.2013, leg. Pešić & Smit, dissected and slide mounted.
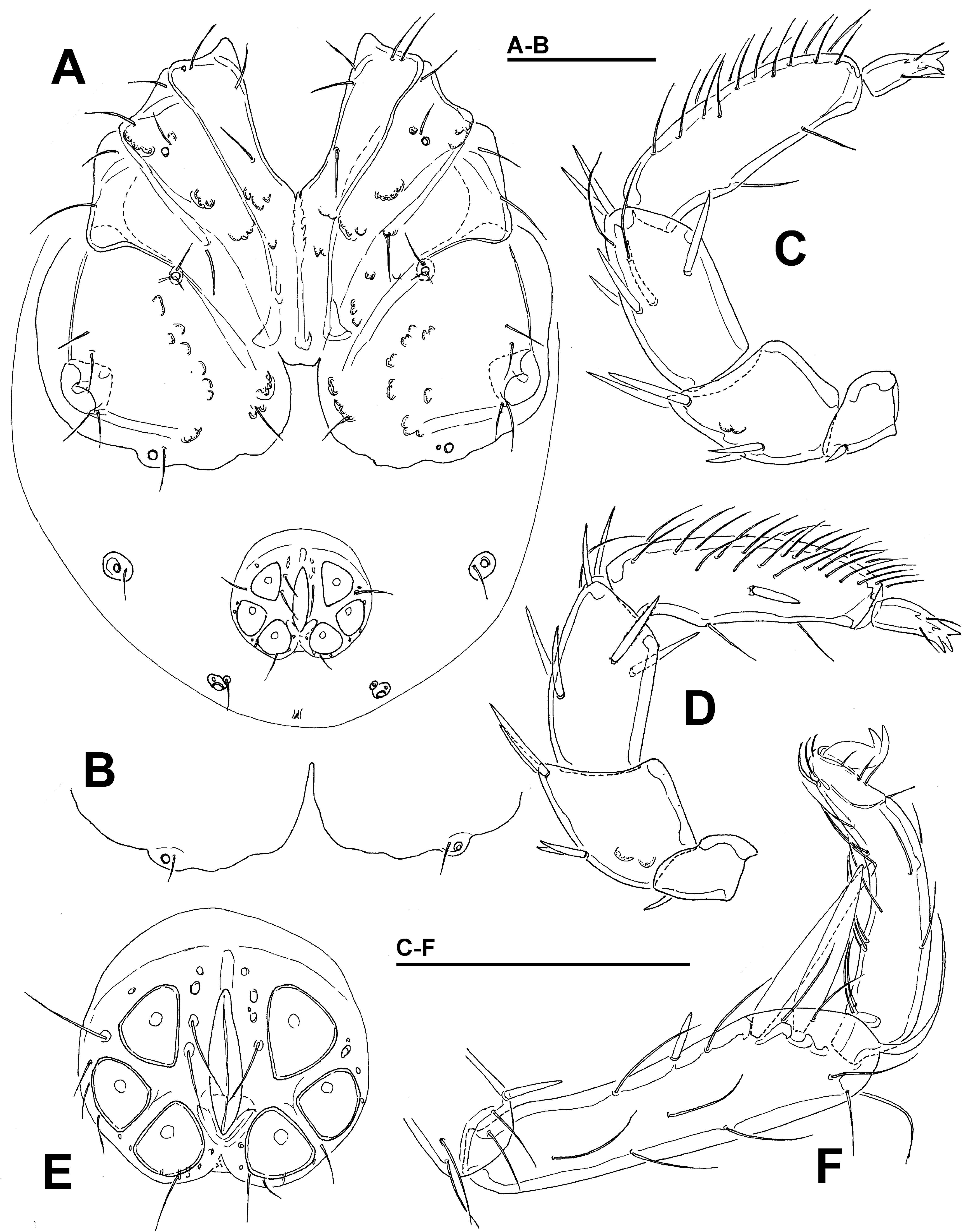
Diagnosis — Female (Male unknown). Integument striated; muscle insertions unsclerotized (visible only as granulated areas); excretory pore smooth; Vgl-1 not fused to Vgl-2; acetabula in a triangular position; P-2 with a pointed ventrodistal protrusion; I-L-5/-6 slightly modified with short and bluntly pointed S-1/-2.
Description — Female. Integument dorsally striated, muscle insertions unsclerotized, glandularia small, round (maximum diameter Dgl 22 µm). Coxal field slender and longish, mediocaudal margin of Cx-I tongue-shaped (Figure 4B). Genital field: gonopore very long, genital plates short and stout flanking the postgenital sclerite, directed laterally, acetabula in triangular position (Figure 4C). Excretory pore smooth; Vgl-1 not fused to Vgl-2. P-2 ventral margin straight, ending in a pointed ventrodistal protrusion, ventral margin of P-3 straight, P-4 slender, insertions of ventral setae flanked by denticles, sword seta fine on the level of distoventral seta (Figures 4E-F). I-L-5 with S-1/-2 slender, with knob-shaped tip, and close to each other; I-L-6 nearly straight, with strong claw (Figure 4D).
Measurements — Idiosoma L 475, W 278; coxal field: L 251; Cx-III W 238; Cx-I+II mL 84, lL 158; genital field L/W 131/116, genital plates L 63, pregenital sclerite W 56, maximum diameter Ac 1-3: 25, 29, 27.
Gnathosoma — palp total L 286; dL/H, dL/H ratio: P-1, 25/22, 1.14; P-2, 64/42, 1.52; P-3, 72/36, 2.0; P-4, 93/20, 4.58; P-5, 32/11, 2.9; L ratio P-2/P-4 0.69. Gnathosomal base vL 75; chelicera total L 163.
Legs — I-L-5 dL 109, vL 83, dL/vL ratio 1.3, maximum H 26, dL/maximum H 4.27, S-1 L 51, L/W ratio 10.8, S-2 L 44, L/W ratio 7.0, distance S-1-2, 3.0, L ratio S-1/2 1.16; I-L-6 L 92, central H 18, L/central H ratio 5.1; L ratio I-L-5/6, 1.19.
Discussion — The new species resembles the hyporheobiontic Atractides pygmaeus (Motaş & Tanasachi, 1948) in many points of view: small dimensions (idiosoma L < 500 μm), unsclerotized excretory pore, slightly modified I-L-5/-6 with short and bluntly pointed S-1/-2 and the presence of well pronounced denticles accompanying the ventral setae of P-4. Atractides pygmaeus differs in having sclerotized muscle attachments, and fused Vgl-1/2, P-2 forming an acute distoventral angle, without projection and acetabula arranged in a weakly curved line (see Gerecke et al. 2016). These clear cut differences, together with the fact that in the male no particular sexual dimorphism is to be expected, justifies the description of the new species based on a single female specimen only.
Etymology — Named after Manas, the heroic warrior featured in one of the longest epic poems in the history of the world “The Epic of Manas” (Kyrgyz: Манас дастаны).
Distribution — Kyrgyzstan; known only from the locus typicus (Figure 10F).
ZOOBANK: 51EA2777-8191-4C77-876B-0ACE5B269136 ![]()





Type material — Holotype male (RMNH), Kyrgyzstan, KR3 Ala Archa NP, rheocrenic spring and first order stream, 42°36.202'N, 074°28.837'E, 1899 m asl., 9.vii.2013, leg. Pešić & Smit, dissected and slide mounted. Paratypes: 12/27/3, same data as holotype, 2/1/1, dissected and slide mounted.
Diagnosis — Coxae without sexual dimorphism (in both sexes Cx-I+II close to Cx-III+IV at their touching edges, but not forming a coxal shield, Vgl-3 not fused to Cx-IV); P-2 ventral margin convex, without distal projection, P-3 with one seta each on medial and lateral surface extending beyond ventral margin of segment; in male S-1-2 separation 19-21 μm.
Description — Adults: General features — Integument striated, muscle insertions unsclerotized. Excretory pore smooth; Vgl-1 not fused to Vgl-2. Palp with weak sexual dimorphism, P-2 ventral margin convex, without projection, P-3 with one seta each on medial and lateral surface, P-4 sword seta between ventral setae. I-L-5 with S-1 and -2 separated, S-1 longish and blunt, S-2 pointed and thicker than S-1, I-L-6 curved, basally thickened, from the centre to the claw furrow with parallel dorsal and ventral margins (Figure 5A). Male: Genital plate anterior margin equally convex, with a narrow border of secondary sclerotization, posterior margin indented, acetabula in an obtuse triangle (Figure 5E); P-2 ventrodistally convex, P-4 thickened, ventral margin slightly projecting near insertion of proximoventral seta (Figures 5C-D). Female. Acetabula in a weakly curved line (Figure 6D); P-2 ventral margin slightly convex, P-4 slender than in male (Figure 6C).
Deutonymph: Provisional genital field with two pairs of acetabula (Figure 7C); P-2 ventral margin convex, P-3 without seta on medial and lateral surface.
Measurements — Male (holotype; in parentheses paratypes, n = 2) — Idiosoma L 628 (631-656), W 416 (425-475); maximum diameter Dgl 26 (27-28); coxal field: L 328 (339-359), Cx-III W 347 (344-375), mL 113 (113-116), lL 204 (216); genital field L/W 117 (120-128)/123 (127-134), maximum diameter Ac 1-3: 38 (41-45), 39 (38-46), 40 (39-46); ejaculatory complex L 111 (125).
Gnathosoma — palp total L 307 (304-317); dL/H, dL/H ratio: P-1, 33/28, 1.17 (31-34/28-30, 1.1-1.2); P-2, 66/47, 1.4 (66/48, 1.36-1.37); P-3, 77/41, 1.89 (75-77/41, 1.85-1.89); P-4, 97/27, 3.6 (101-106/25, 4.0-4.2); P-5, 34/12, 2.9 (31-34/13, 2.5-2.7); L ratio P-2/P-4 0.68 (0.63-0.65). Gnathosomal base vL (106-111); chelicera total L (212-213).
Legs — I-L-5 dL 184 (186-192), vL 123 (119-126), dL/vL ratio 1.5 (1.53-1.56), maximum H 47 (47-52), dL/maximum H 3.9 (3.7-4.0), S-1 L 86 (89-91), L/W ratio 9.2 (9.0-9.5), S-2 L 72 (75-78), L/W ratio 6.2 (5.7-6.2) , distance S-1-2 19 (20-21), L ratio S-1/2 1.19 (1.17-1.19 ); I-L-6 L 129 (128-130), central H 22 (20-21), L/central H ratio 5.9 (6.1-6.4); L ratio I-L-5/6 1.43 (1.45-1.48).
Female (paratype, n = 1) — Idiosoma L 781, W 578; maximum diameter Dgl 27; coxal field: L 403; Cx-III W 456; Cx-I+II mL 141, lL 267; genital field L/W 182/180, genital plates L 130-136, pregenital sclerite W 81, maximum diameter Ac 1-3: 45, 42, 44.
Gnathosoma — palp total L 391; dL/H, dL/H ratio: P-1, 37/34, 1.07; P-2, 84/56, 1.5; P-3, 105/46, 2.28; P-4, 125/30, 4.2; P-5, 42/16, 2.7; L ratio P-2/P-4 0.68. Gnathosomal base vL 128; chelicera total L 266.
Legs — I-L-5 dL 231, vL 148, dL/vL ratio 1.57, maximum H 59, dL/maximum H 3.9, S-1 L 113, L/W ratio 10.6, S-2 L 89, L/W ratio 6.8, distance S-1-2 31, L ratio S-1/2 1.26; I-L-6 L 148, central H 23, L/central H ratio 6.3; L ratio I-L-5/6, 1.56.
Deutonymph (n = 1) — Idiosoma L 409, W 298; provisionally genital field L/W 63/80, ratio 0.79.
Palp: total L 219; dL/H, dL/H ratio: P-1, 23/22, 1.1; P-2, 45/34, 1.35; P-3, 56/28, 2.0; P-4, 69/19, 3.7; P-5, 26/9, 2.75; L ratio P-2/P-4 0.66.
Legs — I-L-5 dL 144, vL 97, dL/vL ratio 1.49, maximum H 41, dL/maximum H 3.55, S-1 L 69, L/W ratio 7.3, S-2 L 63, L/W ratio 5.7, distance S-1-2 13, L ratio S-1/2 1.1; I-L-6 L 100, central H 19, L/central H ratio 5.3; L ratio I-L-5/6, 1.44.
Discussion — Due to a similar shape of palp and I-L-5 and -6, the new species is closely related to Atractides sonkulensis n. sp. (see below) and differs from all other species in having one seta each on medial and lateral surface of P-3. Atractides sonkulensis n. sp. differs in the male in generally smaller dimensions of idiosoma, I-L-5/6 and palps (e.g., idiosoma L < 500 μm, -L-5 dL 145, palp total L < 300 μm), a coxal field merged to an unique plate including Vgl-3, a nearly straight ventral margin of P-2 and a shorter S-1/2 interspace (9-10 μm). In the female A. sonkulensis n. sp. differs in a smaller dimensions of medial suture line of Cx-I+II, genital plates and acetabula, and a relatively longer I-L-6 (L ratio I-L-5/6 1.4).
Etymology — Named after the National Park where the new species was found.
Distribution — Kyrgyzstan; known only from the locus typicus (Figure 10A).
ZOOBANK: CC68E207-1755-4B42-BF73-6E2F3F99C84F ![]()



Type material — Holotype male (RMNH), Kyrgyzstan, KR32 stream near Son Kul Lake, 41°55.480'N, 75°11.865'E, 3066 m asl., 16.vii.2013, leg. Pešić & Smit, dissected and slide mounted. Paratypes: 10/27/0, same data as holotype, 2/1/0, dissected and slide mounted.
Further records — Kyrgyzstan, KR31 River near Son Kul Lake, 41°51.549'N, 75°20.133'E, 3049 m asl., 16.vii.2013, leg. Pešić & Smit 0/3/0; KR34 Son Kul Lake area, rheohelocrenic spring 1, 41°56.236'N, 75°12.686'E, 3093 m asl., 16.vii.2013, Pešić & Smit 0/2/0.
Diagnosis — Coxae with sexual dimorphism (in male coxal field merged to an unique plate, including Vgl-3; in female coxal plates separate); ventral margin of P-2 nearly straight, P-3 with one seta each on medial and lateral surface extending beyond ventral margin of segment; in male S-1/-2 close to each other (separation 9-10 μm).


Description — General features — Integument dorsally striated, muscle insertions unsclerotized. Coxae with sexual dimorphism. Excretory pore smooth; Vgl-1 separate from Vgl-2. Palp with weak sexual dimorphism, ventral margin of P-2 nearly straight, forming a right distal angle, not protruding, P-3 with one seta each on medial and lateral surface, P-4 sword seta between ventral setae. I-L-5: S-1 longish and blunt, S-2 pointed and thicker than S-1, I-L-6 curved, basally thickened. Male: Coxae with extended secondary sclerotization, forming a coxal shield, including Vgl-3 (Figure 8A); genital plate with extended convex margin of secondary sclerotization, posterior margin indented, acetabula in an obtuse triangle (Figure 8E); P-4 thickened, slightly elevated near insertion of proximoventral seta (Figures 8C-D). Female. Coxal plates separate (Figure 9A); acetabula in a weakly curved line (Figure 9B); P-4 more slender (Figures 9C-D).
Measurements — Male (holotype; in parentheses paratypes, n = 2) — Idiosoma L (478-491), W (350-353); maximum diameter Dgl 21 (22-23); coxal field: L 298 (302-322); Cx-III W 291 (288-311); genital field L/W 94/100 (86-88/92-95), maximum diameter Ac 1-3: 31 (24-27), 28 (22-23), 26 (24-23); ejaculatory complex L (105).
Gnathosoma — palp total L 276 (286); dL/H, dL/H ratio: P-1, 27/25, 1.06 (27/27, 1.0); P-2, 58/42, 1.38 (63/42, 1.48); P-3, 67/34, 1.95 (69/36, 1.92); P-4, 94/22, 4.28 (97/23, 4.2); P-5, 30/12, 2.6 (30/12, 2.5); L ratio P-2/P-4 0.62 (0.65). Gnathosomal base vL 89 (102); chelicera total L 161 (170).
Legs — I-L-5 dL 145 (145), vL 98 (101-103), dL/vL ratio 1.48 (1.4-1.43), maximum H 39 (38-40), dL/maximum H 3.7 (3.7-3.9), S-1 L 70 (70), L/W ratio 9.0 (8.1), S-2 L 63 (63-64), L/W ratio 6.7 (6.2-6.3), distance S-1-2 10 (9-10), L ratio S-1/2 1.1 (1.1); I-L-6 L 105 (106-108), central H 19 (20), L/central H ratio 5.4 (5.2-5.4); L ratio I-L-5/6, 1.38 (1.35-1.37).
Female (paratype, n = 1) — Idiosoma L 750, W 570; maximum diameter Dgl 22; coxal field: L 369; Cx-III W 441; Cx-I+II mL 125, lL 242; genital field L/W 159/175, genital plates L 109-114, pregenital sclerite W 79, maximum diameter Ac 1-3: 39, 34, 35.
Gnathosoma — palp total L 359; dL/H, dL/H ratio: P-1, 34/33, 1.04; P-2, 73/50, 1.47; P-3, 98/41, 2.4; P-4, 116/27, 4.35; P-5, 38/14, 2.66; L ratio P-2/P-4 0.63. Gnathosomal base vL 131; chelicera total L 248.
Legs — I-L-5 dL 213, vL 141, dL/vL ratio 1.52, maximum H 50, dL/maximum H 4.3, S-1 L 105, L/W ratio 11.1, S-2 L 75, L/W ratio 6.4, distance S-1-2 27, L ratio S-1/2 1.4; I-L-6 L 150, central H 22, L/central H ratio 6.7; L ratio I-L-5/6, 1.42.
Discussion — See discussion for Atractides alaarchaensis n. sp.
Etymology — Named after Son Kul lake in the vicinity of which the new species was found.
Distribution — Kyrgyzstan; known only from streams near Son Kul lake (Figures 10B, H).
Daday E. von 1903 — Mikroskopische Süßwasserthiere aus Turkestan — Zool. Jb. Syst., 19: 469-553.
Gerecke R. 2003 — Water mites of the genus Atractides Koch, 1837 (Acari: Parasitengona: Hygrobatidae) in the western Palaearctic region: A revision — Zool. J. Linn. Soc., 138(2-3): 141-378. doi:10.1046/j.1096-3642.06-0.00051.x ![]()
Gerecke R., Gledhill T., Pešić V., Smit H. 2016 — Chelicerata: Acari III — In: Gerecke R, ed. Süßwasserfauna von Mitteleuropa, Bd. 7/2-3. Springer-Verlag Berlin, Heidelberg, pp. 1-429.
Jankowskaja A.I. 1972 — Phreatic and spring fauna of Northern Tien Shan and Southern Kisilkum — Trudy Zoologitscheskogo Instituta, Akademiia Nauk SSSR, Leningrad, 51: 5-56. [in Russian]
Lundblad O. 1969 — Indische Wassermilben, hauptsächlich von Hinterindien — Ark. Zool., 22: 289-443.
Pesic V., Smit H. 2009 — New records of water mites of the genus Atractides Koch, 1837 (Acari: Hydrachnidia, Hygrobatidae) from Thailand, Malaysia and Sulawesi (Indonesia), with the description of four new species — Zootaxa, 2240: 1-30.
Pešić V., Smit H. 2014 — Torrenticolid water mites (Acari: Hydrachnidia: Torrenticolidae) from Malaysian Borneo — Zootaxa, 3840(1): 1-72. doi:10.11646/zootaxa.3840.1.1 ![]()
Pešić V., Smit H. 2017 — Neumania kyrgyzica n. sp. a new water mite from Kyrgyzstan based on morphological and molecular data (Acari, Hydrachnidia: Unionicolidae) — Syst. Appl. Acarol., 22(6): 885-894.
Pešić V., Chatterjee T., Das M., Bordoloi S. 2012 — Two rare water mite species (Acari, Hydrachnidia) from the streams of the Indian eastern Himalayan region — Syst. Appl. Acarol., 17(4): 458-464. doi:10.11158/saa.17.4.15 ![]()
Pešić V., Semenchenko K.A., Lee W. 2015 — Further studies on water mites from Korea, with description of two new species (Acari, Hydrachnidia) — ZooKeys, 507: 1-24. doi:10.3897/zookeys.507.9973 ![]()
Ruzskij M.D 1929 — Über einige Arten von Milben (Acarina) am Kurorte ``See Karatschy" — Omskij medicinskij zhurnal, 3-4: 146-149.
Semenchenko K.A. 2016 — New water mites species (Acariformes: Hydrachnidia) from interstitial waters of the Russian Far East — Zootaxa, 4097(4): 545-556. doi:10.11646/zootaxa.4097.4.6 ![]()
Smit H. 2016 — A new hydryphantid genus from the Tien Shan Mountains of Kazakhstan (Acari, Hydrachnidia: Hydryphantidae) — Zootaxa, 4200(2): 324-326. doi:10.11646/zootaxa.4200.2.6 ![]()
Smit H., Pešić V. 2014 — Water mites from Mount Kinabalu and the Crocker Range, Borneo, Malaysia (Acari: Hydrachnidia), with the description of 34 new species — Zootaxa, 3876(1): 1-71. doi:10.11646/zootaxa.3876.1.1 ![]()
Smit H., Gerecke R., Pešić V., Gledhill T. 2015 — On the taxonomic state of water mite taxa (Acari: Hydrachnidia) described from the Palaearctic, part 3, Hygrobatoidea and Arrenuroidea, with new faunistic data — Zootaxa, 3981(4): 542-552. doi:10.11646/zootaxa.3981.4.5 ![]()
Sokolow I. 1926 — Zwei neue Hydracarinenarten aus der Gattung Atractides C.L.Koch — Zool. Anz., 68: 72-76.
Sokolow I.I. 1928 — Zur Kenntnis der Hydracarinenfauna von Buchara — Zool. Jb. Syst, 54(5-6): 467-486.
Sokolow I.I. 1935 — Zur Kenntnis der Hydracarinenfauna der Seen der Katunschen Alpen (Altai) — Issledovaniya Ozer SSSR, 8: 269-273. [in Russian]
Sokolow I.I. 1940 — Hydracarina (1re partie: Hydrachnellae) — Faune de l'URSS. Arachnides, 5(2). Edition de l' Academie des Sciences de l' URSS, Moscou–Leningrad, 24 + 511 pp. [in Russian]
Sokolow I.I. 1948 — Hydracarinen aus Tadjikistan — Denkschriften Akademiker S.A. Sernow: 242-262.
Thor S. 1909 — Beiträge zur Kenntnis der Fauna Turkestans auf Grund des von D.D. Pedaschenko gesammelten Materials. VI. Acarina aus Turkestan und aus dem Distrikte Akmolinsk — Trav. Soc. Natural., Pétersbourg, 39(2): 77-99.
Thor S. 1930 — Einige Acarina, besonders Hydracarina aus Turkestan. Mit dem Nachweis, daß Hjartdalia nicht (wie von C. Walter behauptet) mit Kongsbergia identisch ist — Zool. Anz., 88(7-8): 179-198.
Tjutenkow S.K. 1956 — Hydrobiologische Skizze des Kurgaldshin-Sees — Sbornik Rabot po Ichtiologii i Gidrobiologii, Akad. N. Kasachskoj SSR Institut Zoologii, 1: 124-154. [in Russian]
Tuzovskij P.V. 1987 — New water mites species of the genus Wandesia (Hydryphantidae, Acariformes) from Magadanskaya Province — Biol. Nauki, 3: 36-44. [in Russian, with English summary]
Tuzovskij P.V. 2003 — Two new species of water mites of the genus Sperchon Kramer (Acariformes, Sperchontidae) from Central Asia — Acarina, 11: 215-219
Wiles P.R. 1991 — Rheophilic watermites (Acari: Hydrachnidia) from mainland Malaysia — Acarologia, 32: 41-56.
Wiles P.R. 1997 — Asian and Oriental Torrenticolidae Piersig, 1902 (Acari: Hydrachnidia: Lebertioidea): a revision of the family and description of new species of Torrenticola Piersig and Pseudotorrenticola Walter, from Southeast Asia — J. Nat. Hist., 31: 191-236. doi:10.1080/00222939700770121 ![]()



2017-05-22
Date accepted:
2017-08-18
Date published:
2018-01-31
Edited by:
Kreiter, Serge

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Pešić, Vladimir and Smit, Harry
 Download article Download low definition
Download article Download low definitionDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)