



Two Eocene species of peacock mites (Acari: Tetranychoidea: Tuckerellidae)
Sidorchuk, Ekaterina A.1 and Khaustov, Alexander A.2
1✉ Paleontological Institute, Russian Academy of Sciences, Moscow, Russia.
2Tyumen State University, Tyumen, Russia.
2018 - Volume: 58 Issue: 1 pages: 99-115
https://doi.org/10.24349/acarologia/20184228ZooBank LSID: 086E8B1E-6B45-4250-B67F-9A6C470BFA09
Keywords
Abstract
Plant-feeding trombidiform mites – gall and rust mites, tetranychoid, some tydeoid, eupodoid and heterostigmatic mites – are presently ubiquitous on higher plants. Phytophagy was repeatedly acquired by different lineages of Trombidiformes, and some of these acquisitions might have happened as early as in the Mesozoic (Lindquist 1998). There is, however, very little direct evidence of this evolution, for the fossil record of phytophagous mites is extremely scant. The oldest Tetrapodili, four species of Triasacaroidea, are described from the Triassic (ca 230 mya) amber of South Dolomites in Italy (Schmidt et al. 2012; Sidorchuk et al. 2014). A middle/late Eocene (ca 35 mya) Eriophyidae specimen from Maslin fossil beds in South Australia is described as a species in a Recent genus, Aculops Keifer, 1966 (Southcott and Lange 1971). Two possible spider mite species are described from Eocene (56 to 33 mya) Baltic amber by C.L. Koch in Koch and Berendt (1854): Panonychus gibbus (C.L. Koch, 1854) and Schizotetranychus? brevipes (C.L. Koch, 1854), but their morphology is only partly known (Oudemans 1937). One species of a peacock mite (Tetranychoidea: Tuckerellidae), Tuckerella fossilibus Khaustov, Sergeyenko, Perkovsky, 2014, is known from Eocene Ukrainian (Rovno) amber. It was described from a single poorly preserved specimen. A close preparation was not undertaken due to the risks involved, and although this resulted in its description being incomplete, there was not doubt as to the species identity. Recently, a new peacock mite fossil was found in Eocene Baltic amber by Thomas Weiterschan, who has kindly lent the specimen for our study. The mite, fossilized in a defensive posture characteristic for extant tuckerellids, appeared to represent a distinct species, new to science.
By this contribution, we aim to describe the new species, Tuckerella weiterschani n. sp., and augment the description of T. fossilibus after closer examination of its holotype.
The Baltic amber piece with a single inclusion, representing a new species of Tuckerella, originally in the collection of Thomas Weiterschan (TW), collection identification number TW 1288. The piece is transferred to the collection of the Geological-Paleontological Museum and University of Hamburg (GPIH), and assigned a new collection number GPIH 4598. The piece is translucent, light yellow in colour. The fragment measuring 1x0.7x0.7 mm, containing the mite, was extracted and polished on six sides. For a review, and further data on the geology, of Baltic amber, see Weitschat and Wichard (2002). The Holotype of Tuckerella fossilibus is a Rovno amber piece, found in Klesov (Fedorovka) locality in Ukraine. The geology and fauna of this locality is reviewed by Perkovsky et al. (2010). The specimen is deposited in I.I. Schmalhausen Institute of Zoology, National Academy of Science, Kiev (SIZK), collection number KF-2237. A fragment, sized 2x1x0.5 mm, of the piece was mounted in the Buehler EpoThin epoxy resin between two 1 cm round coverslips. The fragment has a fracture that runs through the anterior part of the mite specimen. The preparation is labeled and glued with a hot-melt adhesive to a 1.5 mm-thick transparent plastic frame with a round aperture, allowing for observation from dorsal and ventral sides. The frame was also labeled.
Amber pieces were cut and polished following protocols of Sidorchuk (2013), with tools described by Sidorchuk and Vorontsov (2016).
All observations were done with light microscopy. Image stacks were obtained with Zeiss AxioImager A2 compound microscope with dry lenses and differential interference contrast (DIC) illumination with AxioCam 506 color digital camera. Bright field and polarized light micrographs were obtained with Nikon Eclipse 800 microscopes with 60x water immersion lens, Nikon DS-Fi1 and Nikon D7000 digital cameras. All images were corrected for light, color, noise and sharpness using Adobe Lightroom, then processed with Helicon Focus algorithm A, always with some detail manually transferred from the individual focal planes to the resulting images. These images are further referred to as ``layered''. Original image stacks are available through Figshare, doi:10.6084/m9.figshare.c.3743399 ![]() .
.
Drawings are made in Adobe Photoshop with stacks of the light micrographs as a background or in InkScape with a live capture from the Amscope MU900 camera mounted on Nikon Eclipse 800 microscope as a background (Sidorchuk and Vorontsov 2014).
The terminology is that developed by F. Grandjean for acariform mites (see Travé and Vachon 1975 for review and bibliography, and Lindquist 1985 for application to Tetranychoidea); shapes of setae are described using botanical glossary by Beentje (2010). Idiosomal setae designations follow Beard et al. (2013), i.e. prodorsal setae are named sc 1, sc 2, v 1, v 2, setal row D contains 6 setae and row E contains 3 setae. There are three versions of the homologization of tuckerellid tarsal setae to those of the other acariform mites: (1) Quiros and Baker 1984, (2) Lindquist 1985, reproduced in Servin and Otero 1989, and (3) Bochkov et al. 2009. We accept here Lindquist's version, in which the primilateral setal pair (pl) is considered present in the larva of Tetranychoidea (including Tuckerellidae), the tectal setal pair (tc) and seta l 1 " are added in postlarval stages, dorsal seta d and antelateral setal pair (a) are absent. Leg setal counts are written from trochanter to tarsus as the number of tactile/eupathidial setae followed by the number of solenidia in parentheses. Leg setation formulae represent the cumulative data from both legs. For example, when on the right tibia III the setae l' and v' are visible, while on the left these are d and v', observed setal count is d+l'+v'=3 setae.
Comparative data on the leg setation was obtained from the following species and works: T. pavoniformis Ewing, 1922 and T. ornata Tucker, 1926 – from Baker and Pritchard 1953; T. litoralis Collyer, 1969, T. japonica Ehara, 1975, T. anommata Smith Meyer and Ueckermann, 1997, T. channabasavannai Mallik and Kumar 1992, T. revelata Beard and Walter, 2005 – from the original descriptions of these species. Hypotheses on the setal homologies are ours, but for the latter species Jennifer Beard has generously supplied additional sketches with her setal homologies, confirming our hypotheses. In addition, Evert E. Lindquist (EEL) has kindly examined and sketched for us tarsi I and tarsus II of a male T. flabellifera Miller, 1964 and tarsus I of female T. ornata, kept at the Canadian National Collection of Insects, Arachnids and Nematodes, Ottawa); for these, setal homologies are by EEL.
The measurements were taken from edge to edge of a sclerite or seta in its widest part, choosing where possible the structures oriented perpendicularly to the optical axis of the microscope. As the orientations were non-ideal, all measurements are to be treated as the minimal estimates, and are rounded to closest 5 if over 50. The unit is micrometer (μm).
Order Trombidiformes Reuter, 1909
Family Tuckerellidae Baker & Pritchard, 1953
Genus Tuckerella Womersley, 1940
Type species: Tenuipalpus ornatus Tucker, 1926, by original designation
ZOOBANK: AB714EE7-5602-4792-A9A0-B42618B1E983 ![]()
Description — Tritonymph or adult, gender unknown. Length of idiosoma – 275, width – 155, height – 55; body length including gnathosoma – 310.
Gnathosoma (Figs 1, 2, 4) — Only right palp clearly visible. Palpgenu dorsally with seta d, reaching just beyond tip of tibia. Palptibia with large, hooked tibial claw and three setae: d, slightly longer than palpgenu; short l", reaching to tip of tibial claw; medium-sized l', about as long as palptibia+tibial claw. Palptarsus completely covered by tibial claw in dorsal view, and elongate, roughly necktie-shaped as visible in lateral view, 14 long, phaneres not visible. Subcapitulum long and narrow, with alveolus of seta m visible basally and one pair of small peg-like adoral setae visible apically (Fig. 2, arrow). Peritremes prominent and emergent (Figs 1H, 4A). Stylophore and cheliceral stylets not visible.








Idiosomal dorsum (Figs 1, 3, 4A) — Visible ornamentation reticulate (Fig. 1D–F). Prodorsum with 4 pairs of setae (vertical v 1, v 2, scapular sc 1, sc 2). Setae v 2, sc 1 and sc 2 fan-like (Figs 1D, 4A); setae v 1 broadly lanceolate, with tapered tip (Fig. 1H). Two pairs of eyes located anterolaterally to bases of setae sc 1 (Figs 1F, 4A, oc); posterior eye distinctly larger than anterior. Hysterosoma with 7 setae in C-row, 6 in D-row, 3 in E-row, 2 in F-row, 8 in H-row. All setae in rows C–F broadly obovate to orbicular. Setae c 1–c 3 and d 1–d 3 of medium size (12x20), subequal. Setae e 1 and e 2 slightly smaller than c 1–c 3 and d 1–d 3. Setae f 1 and f 2 (8x14) distinctly smaller than e 1 and e 2; f 2 situated anterolaterally to bases of f 1. Setae c 4 (20x30) distinctly larger than c 1–c 3; c 5 (20x40) more elongate and narrow than c 4; setae c 6 and c 7 similar to c 5, but with stronger tapered tips, especially well visible on left side of body (Figs 1D, 3). Setae d 4–d 6 and e 3 similar in shape to c 4–c 7, but clearly larger (d 6 30x60). Distinct apophyses of h-series setae situated close to each other in one transverse line. Setae h 1 short, narrowly lanceolate, barbed, 30 long (Fig. 1G), much shorter than filiform and weakly barbed setae h 2–h 8, . Setae h 2–h 8 longer than body (h 2 440) and directed dorsad and anteriad (Fig 1A, C, G, J). Only shield margins visible are anterior margins of dorsal shields C and D (Figs 1A, 3).
Idiosomal venter (Fig. 1B) — Somewhat collapsed, mostly obscured by legs. Cuticle lightly striate, setation not discernible. As far as visible, typical for females of Recent Tuckerella species (see for example Fig. 2 in Beard and Walter 2005). Genital and anal openings discernible, but no setae visible with certainty.
Legs (Figs 1B, J; 4) — Leg I (Fig. 4A, A'). Femur with 3 large, fan-like dorsal and lateral setae (d, l', l"), and 2 small, setiform ventral setae visible (probably ventral v" and v 1 "). Genu with 5 large, fan-like dorsal and lateral setae (d, l', l 1 ', l", l 1 ") and one visible setiform seta v". Tibia with large, fan-like setae l", l 1 ", l'; and more narrow d; solenidion φ baculiform, situated on short protuberance; single visible seta v" small, setiform. Tarsus with two claws and empodium with tenent hairs (Figs 1A, 4A, A'); two solenidia present, ω1 about 1.5 times longer than ω2; unguinal setae (u) poorly visible, only on left leg, modified, comb-like; other setae on tubercles: abaxial members (") of fastigial (ft) and primilateral (pl) pairs long, distinctly longer than tarsus, their adaxial members ft' and pl' about half as long; pl' weakly barbed; tectal (tc) smooth, tapered; proral p" blunt, thick, probably eupathidial, possible alveolus of p' visible on right leg; simple primiventral seta pv visible on left tarsus; on right tarsus, alveolus in position l 1 " visible and what may be seta l 1 " discernible. Leg II (Fig. 4B) setation partly visible, tip of tarsus and trochanter hidden in deformed venter. Femur with 3 fan-like – d and (l) – and one poorly visible simple seta, possibly basiventral bv". Genu with 4 fan-like setae (d, (l), l 1 '), tibia with 3 fan-like setae – d, (l) – visible. Tarsus with one blunt phanere on prominent tubercle (probably solenidion ω) and 7 simple setae or their alveoli visible: pv', pairs (p) and (tc) on low tubercles, seta pl" and possible alveolus of ft". Legs III and IV (Fig. 4C, D, D') visible almost entirely, with paired claws, paired empodia and at least 4 pairs of tenent hairs each. Setation: leg III trochanter with single thick, heavily barbed seta l' visible, femur with 2 setae: simple euventral ev' and short, narrow, barbed d; genu with single short, narrow, barbed seta d visible, tibia with 3 setae: short, narrow, barbed d, fan-like l', simple v'; tarsus with two setae and three alveoli visible, setal pair (p) and solenidion apparently absent. Leg IV trochanter and femur without visible setae; genu with setiform v' and short, narrow, barbed d, tibia with single short, narrow, barbed seta d visible. Tarsus with 7 setae: pairs (ft), (tc) and (u) visible dorsally (in ventral aspect of mite, Fig. 4D'), seta pv' – in lateral aspect (Fig. 4D). Observed leg setation formulas: leg I: ?-5-6-5(1)-12(2); leg II: ?-4-4-3-7(1); leg III: 1-2-1-3-5; leg IV: ?-?-2-1-7.
Holotype and type repository — Originally from private collection of Thomas Weiterschan, coll. TW 1288, housed in the Geological-Paleontological Museum and University Hamburg (GPIH), collection number GPIH 4598. The holotype is the only inclusion in a rectangular light yellow amber piece measuring 1x0.7x0.7 mm, polished on six sides, stored in plastic O-ring-capped vial with water and preservative Thymol (Fig. 1A).
Type locality, occurrence and geological age — Type locality is Baltic Sea Coast, amber in Blue Earth fossil beds, dated late Eocene. Only known from the type specimen and locality.
Etymology — The new species is named in honor of Thomas Weiterschan, who discovered the specimen, promoted its study and assured its holotype deposition in a public repository.
Remark — The holotype and only known specimen of T. weiterschani has emergent peritremata, leg I tarsal setae l 1 " and leg I genual and tibial setae (l 1) present. These features suggest that the specimen is at least tritonymphal (Servin and Otero 1989, Beard and Walter 2005). In the presence of the tapered setae v 1 and lateral idiosomal setae and in the absence of clear view on the genital area, however, we cannot distinguish between an adult and a tritonymphal stage. Emergent peritremes in immature tuckerellid mites, not mentioned in the literature to our best knowledge, do occur at least in four Recent species starting from the deutonymphal stage (J. Beard, pers. comm.), and are thus inconclusive. The Diagnosis below is hence preliminary, and only uses those characters known to be relatively stable through the last two stages of the tuckerellid ontogeny.
Differential diagnosis — By the presence of seven pairs of subequal filiform setae h 2–h 8 the new species is similar to five Recent species: T. coleogynis Jorgensen, 1967, T. elegans Rossi de Simons, 1972, T. eloisae Servin and Otero, 1989, T. hypoterra McDaniel and Morihara in McDaniel et al. 1975 and T. spechtae Womersley, 1957. The new species differs from T. spechtae and T. hypoterra by the relative positions of setal pairs f 1 and f 2, f 1 being inserted clearly posterior to f 2 in T. weiterschani, as opposed to slightly anterior in these two species; in addition, T. spechtae has setae h 6 shorter than the other flagellate setae. From T. eloisae the new species differs by its much smaller size (total body length – including gnathosoma – 310 in T. weiterschani vs. 480 in tritonymph, 473 – 566 in adults of T. eloisae). T. elegans, only slightly larger than T. weiterschani (adult idiosomal length 342 – 370 in T. elegans vs. 275 in T. weiterschani), and T. coleogynis differs from the new species by its setae v 1 having a smooth edge and uneven reticulation vs. these setae widely lancetiform, with tapered tips in T. weiterschani (the validity of this character is only certain if T. weiterchani holotype is an adult female), and having setae h 2–h 8 as long as body, vs. at least 1.5 times body length in T. weiterschani. T. coleogynis, whose size is unknown, is the most similar to T. weiterschani, differing in the shape of dorsolateral idiosomal setae (only significant if T. weiterchani holotype is an adult female) and distribution of setulae on the setae h 2–h 8: present only in basal part in T. coleogynis, evenly distributed throughout the length in T. weiterschani.
Tuckerella fossilibus Khaustov, Sergeyenko & Perkovsky, 2014, p. 367, Figures 1–5.
Redescription — Deutonymph. Length of idiosoma – 245, width – 165.
Gnathosoma (Figs 5D, 7B, C) — Left palp covers main part of right palp dorsally, both palps preserved in a way to cover infracapitulum dorsally. Palpgenu dorsally with seta d slightly not reaching base of tibial claw. Palptibia with large, hooked tibial claw and three pairs of setae: d slightly longer than length of palpgenu; short l", reaching to tip of tibial claw; medium-sized l', about length of palptibia+tibial claw. Inner margin of tibial claw transversely striate. Palptarsus with 5 visible phaneres: 3 terminal, blunt-ended (probably palpal eupathidia), one short dorsal and one short ventral. Subcapitulum long and narrow with visible stylet protruding apically (Figs 5D, 7B, st); ventrally with one pair of long subcapitular setae m. Peritremes not evident. Oval stylophore, ca 100x40, visible inside body (Figs 5A, 6, sty).

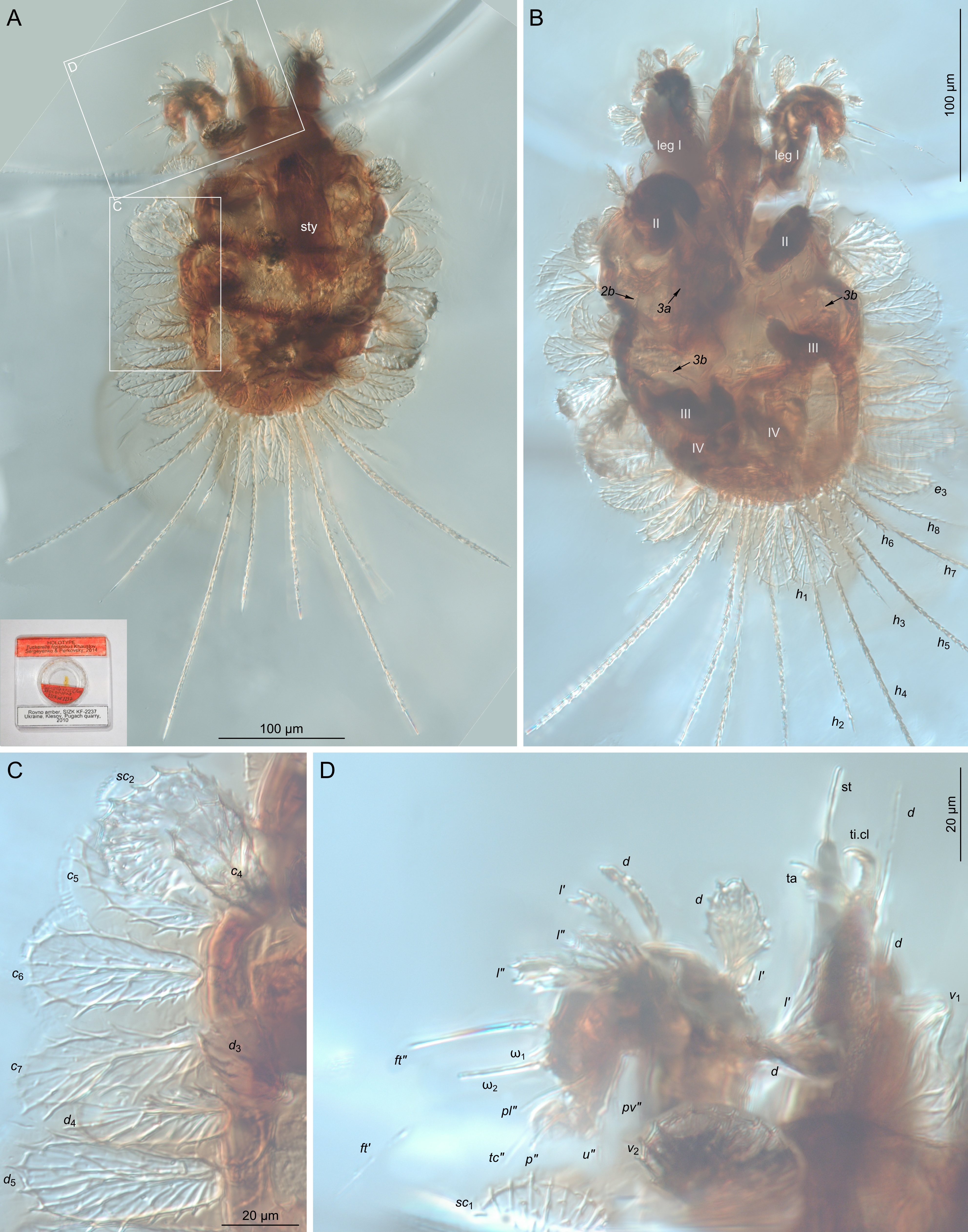




Idiosomal dorsum (Figs 5, 6, 7B) — Cuticular ornamentation, where visible anterodorsally, irregularly reticulate (Fig. 5A). Number of dorsal idiosomal setae as in T. weiterschani n. sp. Setae v 2, sc 1 and sc 2 fan-like (Fig. 5C, D); v 1 considerably different from other prodorsal setae, with smooth margins and weakly tapered tip (Figs. 5D, 7). Two pairs of eyes located anterolaterally to bases of setae sc 1; posterior eye distinctly larger than anterior (Fig. 7B, oc). All setae of C, D, E and F-rows broad, flat, ovate to obovate in shape: c 1-c 3, d 1-d 3 and e 1, e 2 of medium size, subequal (16 – 18 x 24 – 28); f 1 and f 2 slightly smaller than e 1 and e 2; f 2 situated on same transverse level as f 1. Lateral setae c 5-c 7, d 4-d6 and e3 distinctly larger than central, more elongate, with rounded tips, largest d 6 28 x 60. Setae h 1 large, flat, obovate (27 x 55), setae h 2 –h 8 flagellate: h 2, h 4, h 5, h 7 long, barbed, tapered (180, 270, 180, 220 respectively), h 6 short, thick, heavily barbed, blunt (40); h 3, h 8 intermediate in length and thickness, barbed, tapered (80). Only anterior margins of dorsal shields C and D clearly visible (Fig. 6).
Idiosomal venter (Figs 5B, 7A, D, E, F) — Dark and collapsed, obscured by legs. Only coxal setae are partly discernible: 1a-c and 2b smooth, fine, 1a longest; 2c long, thick, heavily barbed. Genital and anal openings indiscernible.
Legs (Fig. 7) — Dark and curled, mostly visible in ventral aspect, with some setae poorly discernible. Leg I (Fig. 7A, B): trochanter without visible setae; femur with three large obovate setae (d, l', l") and setiform bv", other ventral setae not visible. Genu with three large obovate setae (d, l', l") and setiform v", other ventral setae not visible. Tibia with large, obovate seta l" and narrower d, l'; ventral setae not visible; solenidion φ baculiform, situated on short tubercle. Tarsus with two claws and empodium, each with clearly visible tenent hairs; two solenidia present, ω1 about 2 times longer than ω2; setae: (u) simple and short, abaxial ('') members of pairs (ft) and (pl) long, distinctly longer than tarsus, pl' about half shorter on right leg, absent on left leg; (tc) situated on tubercles, fine, tapered, (p) blunt, short, apparently eupathidial, difficult to discern where hidden behind ambulacrum; pv' setiform. Leg II (Fig. 7C, D): trochanter not discernible; femur with long, thick, heavily barbed seta bv" and 3 obovate large setae d and (l) (d and l" artificially split on left leg); genu with 3 obovate setae visible; tibia with 3 obovate and one or two long simple seta visible ((v) on left, only v" on right leg); tarsus with ambulacrum as in leg I (well visible on right leg), one solenidion, and 10 smooth setae: long ft", shorter ft', (pl), (tc), (u) (one member of pair visible on each leg), p" and pv'. Leg III (Fig. 7E, F): trochanter with single visible seta l' thick, long, heavily barbed; femur with small obovate seta d and smooth simple ev'; genu with single visible small obovate seta l'; tibia with 2 clearly visible setae, simple v' and small obovate l' (visible transversally on right leg), and one spot that may be alveolus of seta d; tarsus with ambulacrum like in leg II and 7 simple setae (only 4 clearly visible on right leg): (ft), (tc), (u) and pv'. Leg IV (Fig. 7 H, J): poorly visible, dark, trochanter and setation of femur indiscernible; genu with one well visible narrowly obovate seta d on left leg, no visible setae on right; tibia with 2 ventral setae well visible, two discernible on right leg, one well visible seta and one discernible on left; tarsus with two claws discernible, 3 setae visible on left and one on right leg. See figure for possible setal homologies. Observed setation (trochanter to tarsus, solenidia in parentheses): leg I: ?-5-3-4(1)-11(2); leg II: ?-4-3-5-10(1); leg III: 1-2-1-2/3?-7; leg IV: ?-?-1-2?/4?-?.
Remarks — Khaustov et al. (2014) in original description of T. fossilibus were not able to identify its life stage. Now, after more detailed description we can suggest that it is probably a deutonymph. Our opinion is based on the setation of leg I, which shows: (1) two tarsal solenidia, (2) absence of the tarsal seta l 1 ', (3) three obovate setae on each genu and tibia. According to observations by Quiros-Gonzalez and Baker (1984), Lindquist (1985), Beard and Walter (2005) and Beard et al. 2013, although there is some interspecific variation, the second tarsal solenidion is deutonymphal, tarsal seta l 1 ' is tritonymphal, and three obovate setae on first genu and tibia are only present in deutonymphs: known protonymphs are found to have 1–2 obovate setae per segment, while tritonymphs and adults have five.
Addition to the differential diagnosis — Certain caution is necessary when using this comparison, for the immature specimen is being compared to adults. Based on the available descriptions (Quiros-Gonzalez and Baker 1984, Lindquist 1985, Beard and Walter 2005, Beard et al. 2013), ontogenetic changes of the opisthonotal setation may affect setae v 1 (in T. revelata they are obovate, with rough edge, in larva, with outline smoothing out in the subsequent stages), leaf-shaped C-H-row setae (relatively narrower and more tapered in the immatures) and flagellate H-row setae (becoming relatively shorter during ontogeny). T. fossilibus is so far unique tuckerellid species known to have obovate setae h 1 similar in size to the lateral setae of C-, D- and E rows. By the shape of proterosomal setae v 1 and the relative shortness of setae h 1 and h 6, deutonymphal T. fossilibus is similar to adults of T. pavoniformis (Ewing, 1922), T. indica Prasad, 1973 and T. channabasavannai Malik and Kumar, 1992. It differs from them by much larger setae h 1 and by the differentiated shapes of the other H-row setae (T. pavoniformis has setae h 2-h 5, h 7 and h 8 all similar, whip-like (Baker and Pritchard 1953), as do T. indica and T. channabasavannai). By the shape of setae v 1 T. fossilibus is also similar to T. japonica Ehara, 1975 and to T. kumaonensis Gupta, 1979, but these species have three pairs of comparatively small leaf-like setae in h-series.
Holotype container — Amber fragment with holotype, sized 2x1x0.5 mm, is mounted in the Buehler EpoThin epoxy resin between two 1 cm coverslips. The preparation is glued with a low-temperature hot-melt adhesive to a 1.5 mm-thick transparent plastic frame (Fig. 5A).
With the restricted number of known fossils of the phytophagous mites, the discovery of a tuckerellid specimen in Baltic amber appears lucky. Unlike saprophagous mites, whose recently redescribed species are the same for the two amber deposits (Sidorchuk and Norton 2010; 2011a, b; Sidorchuk and Klimov 2011), the tuckerellids from Rovno and Baltic ambers are not conspecific. Their dissimilarity is indicative of a morphological diversity in Eocene (ca 40 mya) tuckerellids comparable to that found among Recent species. It is difficult, though, to discuss the groups of species in Tuckerellidae, for the family is in much need of a thorough revision, as most species are only cursorily illustrated. We can only attempt some general implications based on the morphological similarities discussed in Remarks, as follows.
Ethological implication: the defensive posture. Holotype of T. weiterschani is captured in the posture, first described as defensive by Ochoa (1989) in Tuckerella knorri Baker and Tuttle, 1975. When disturbed, a tuckerellid mite makes a few quick movements, alternately raising and lowering its `tail' of the h-setae. The whip-like setae of the H-row are often long enough to cover the whole body, legs and gnathosoma of the mite, possibly disturbing an intruder. Our finding shows (Figs 1A–C) the defensive adaptation of a peacock mite dates back at least to the Eocene.
Ecological and geographical implications. Tuckerellidae are suctorial plant feeders, like other tetranychoid mites. Recent species are known to exploit both aboveground and underground parts of the plants, including bark and roots (Jorgensen 1967, McDaniel et al. 1975, Beard et al. 2013). The host range of even an individual species may include multiple plant species, not necessarily closely related: for example, T. japonica is known from Ulmaceae, Theaceae, Actinidiaceae, Ebenaceae, Ericaceae, Rhamnaceae and even coniferous Juniperaceae (Beard et al. 2013). Tuckerellidae are found in climes from humid tropical through Mediterranean and hot desert, but also in cold arid areas. Their presence in amber, therefore, is not informative with regard to the amber forest composition or climate.
Evolutionary implications: setal variation. Our specimens appear to have asymmetrical setation at least on tarsi I (Figs 4, 7, 8): KF-2237 (deutonymph) does not show seta pl' on the left (Figs 7B, 8C), while this seta is present on the right (Figs 7A, 8D), and GPIH 4598 (tritonymph or adult) has at least an alveolus in l 1 " position on the right leg (Figs 4A', 8A), while there is no such seta or alveolus on the left (Figs 4A, 8B). This encouraged us to review what is known on the tuckerellid tarsi (see Material and Methods). Below, we only consider tarsi I, and only the setae (pl), (ft) and l 1 ", for they are the best visible and the least likely overlooked. It appeared that these setae vary within modern tuckerellid species. In T. ornata, seta l 1 " is present in the specimen examined by E. Lindquist, absent in Baker and Pritchard 1953 Fig. 6b. In T. revelata, variable setae appear to be ft' (absent in female, Fig. 4A by Beard and Walter 2005, present in all other illustrated immature and adult legs) and pl' (absent in protonymph, Fig. 12A by Beard and Walter 2005). Note that although Beard and Walter 2005 give setal formulae 14(2) for adult and tritonymph and 13(2) for deutonymph tarsus I in the text, they illustrate 12 setae plus 2 solenidia for the adults and tritonymph and 11(2) for the deutonymph, which is in accordance with the otherwise known maximum complement in this family. The discrepancy is due to the authors' counting for the solenidia twice: (1) in the total count of phaneres and (2) in parentheses (J. Beard, pers. comm., June 2017). Seta pl' also is variable in T. anommata: present according to the counts, illustrated for the male, but lacking in the illustrated female left tarsus, where the remaining seta pl" has an almost dorsal position (Smith Meyer and Ueckermann 1997, Figs. 89 and 85, respectively).


Each of them being admittedly questionable, these observations considered together evoke a long-standing discussion on the possible link between the variation and evolution of leg setae in mites. There are two notions on this link. Based on his 46-year studies on Acariformes, including rearing experiments on parthenogenetic oribatid mite Platynothrus peltifer, Grandjean (1974) formulated the hypothesis of the RVA – regressive evolution (of a seta in the simple row within a lineage) by the alternating vertitions. Unilateral absence of a seta, according to this hypothesis, is an indication of its future disappearance, and the frequency of such absences should grow, albeit very slowly, in a lineage of many generations. Wauthy et al. (1991) brought to acarology another notion, that of the fluctuating asymmetry (Ludwig 1932, Van Valen 1962) – the phenomenon common for bilateral organisms, variation resulting from the random developmental accidents and having no evolutionary implications. In this model, the frequency of absence of a particular seta is not indicative of its future loss, and this parameter for a particular seta is not inheritable. The latter notion is developed and substantiated with new observations and statistical analysis (see Wauthy et al. 1998, Leponce et al. 2001), studying three strains of the haplodiploid prostigmatan Tetranychus urticae, although each with two generations (mothers and sons) only. Although their results were statistically significant, this evolutionary experiment was very far from analyzing `hundreds, maybe hundreds of thousands of generations' that Grandjean (1974) considered necessary for a seta to become lost by alternating vertitions. He (Grandjean, 1974: 779) has actually predicted that no notable change of the vertition frequency would be statistically observable within ten or twenty generations of a parthenogenetic lineage – thus, he indicated that the RVA hypothesis cannot be rejected by a feasibly long artificial experiment.
Paleontological evidence somehow shows us the result of an unfeasibly long evolutionary experiment. Although they are not linked by a direct lineage from mother to daughter, the two fossils described here on one hand and the modern tuckerellids described in the literature on the other hand are random specimens within the same genus, separated by at least 40 million generations. These specimens seem to show variations of the same, eustatic, setae, which may mean that the frequency of their absences, or vertitions, is high in Recent tuckerellids and was already high in the Eocene ones. In the light of RVA, suggesting a slow but steadily augmenting growth of the frequency of vertitions in the course of evolutionary time, presence in the Recent Tuckerella of the setae commonly hit by vertitions 40 million generations ago in the same genus would be extraordinary. In the light of the fluctuating asymmetry notion, such presence does not seem overly surprising: high frequency of the asymmetrical expression of some leg setae may be part of the stability of the tuckerellid habitus itself. Fossils show us that the body plan of Tuckerella was already fully formed forty million years ago, and it apparently kept existing without notable change since that time. This remarkable stasis possibly included the degree of imperfection of the mite body's symmetry. To learn whether this is true or not, and whether this is widespread in Acari, careful studies of leg setations in Recent and fossil mites are necessary.
We are grateful to Thomas Weiterschan, who discovered, showed and loaned to E.S. the mite named in his honor, and to him and Carsten Gröhn for organizing smooth transfer of the specimen to the Hamburg University collection for public repository; to Evgeny Perkovsky for organizing T. fossilibus holotype loan from the SIZK. We cordially thank Evert Lindquist for his generous help, looking for, illustrating and discussing setal homologies of the tuckerellid legs. Thorough review of the manuscript by Jennifer Beard, productive discussion, including her multiple illustrations and sharing of unpublished data are deeply appreciated. Constructive criticisms of the manuscript by two anonymous reviewers are gratefully acknowledged. Collection study by E.S. in Hamburg was supported by Arbeitskreis Bernstein and wonderfully organized by Carsten and Jutta Gröhn, to whom E.S. extends her thankfulness.
Baker E.W., Pritchard A.E. 1953 — The family categories of tetranychoid mites, with a review of the new families Linotetranidae and Tuckerellidae — Ann. Entomol. Soc. Am., 46: 443-258. doi:10.1093/aesa/46.2.243 ![]()
Baker E.W., Tuttle D.M. 1975 — A new species of Tuckerella (Acarina: Tuckerellidae) from Thailand — U. S. Dept. Agr. Coop. Econ. Ins. Rpt., 25(17): 337-340.
Beard J.J., Ochoa R., Childers C.C., Bauchan G.R., Shepard M. 2013 — Travelling with tea: a Tuckerella's tale — Exp. Appl. Acarol., 59(1-2): 177-202. doi:10.1007/s10493-012-9627-9 ![]()
Beard J.J., Walter D.E. 2005 — A new Tuckerella (Prostigmata: Tetranychoidea; Tuckerellidae) from Australia with description of all stages and a discussion of the tritonymphal stage — Acarologia, 45(1): 49-60.
Beentje H. 2010 — The Kew plant glossary. An illustrated dictionary of plant terms — Kew: Royal Botanic Gardens. pp. 164.
Bochkov A. 2009 — A review of mites of the parvorder Eleutherengona (Acariformes: Prostigmata) — permanent parasites of mammals — Voltzit O. V. (Ed) Moscow: Acarina, KMK Scientific Press. pp. 149.
Collyer E. 1969 — Two species of Tuckerella (Acarina: Tuckerellidae from New Zealand — New. Zeal. J. Sci., 12(4): 811-814.
Cuvier G. 1812 — Sur un nouveau rapprochement à établir entre les classes qui composent le Règne animal — Annls Mus. Hist. nat., 19: 73-84.
Ehara S. 1975 — Description of a new species of Tuckerella from Japan (Acarina: Tuckerellidae) — Int. J. Acarology, 1(2): 1-5. doi:10.1080/01647957508683741 ![]()
Ewing H.E. 1922 — Three new species of peculiar and injurious spider mites — Proc. Entomol. Soc. Wash., 24(4): 104-108.
Grandjean F. 1974 — Caractères anormaux et vertitionnels rencontrés dans des clones de Platynothrus peltifer (Koch). Chapitres VII à XIII de la deuxième partie — Acarologia, 15 (1973)(4): 759-780.
Gupta S.K. 1979 — Tuckerella kumaonensis, sp. nov. (Acarina: Tuckerellidae) from India — Orient. Insects, 13(1-2): 229-230. doi:10.1080/00305316.1979.10433560 ![]()
Jorgensen C.D. 1967 — A new species of Tuckerella (Acarina: Tuckerellidae) from Nevada — Entomol. News, 78(6): 141-146.
Keifer H.H. 1966 — Eriophyid studies B-21 — Bureau of Entomology, California Department of Agriculture. pp. 24.
Khaustov A.A., Sergeyenko A.L., Perkovsky E.E. 2014 — First fossil record of mites of the family Tuckerellidae (Acari: Tetranychoidea) from Rovno amber with description of a new species — Int. J. Acarology, 40(5): 367-369. doi:10.1080/01647954.2014.933253 ![]()
Koch C.L. 1936 — Deutschlands Crustaceen, Myriapoden und Arachniden — Regensburg. H. 1. N. 10.
Koch C.L. 1939 — Deutschlands Crustaceen, Myriapoden und Arachniden — Regensburg. H. 173. N. 9.
Koch C.L., Berendt G.C. 1854 — Die im Bernstein befindlichen Crustaceen, Myriopoden, Arachniden und Apteren der Vorwelt — In: Berendt G. C. (Ed) Die in Bernstein befindlichen organischen Reste der Vorwelt, gesammelt, in Verbindung mit Mehreren bearbeitetet und herausgegeben, Berlin: Commission der Nicolaischen Buchhandlung. pp. 103-111; 124; T. XVII, XIII.
Leponce M., Noti M.-I., Bauchau V., Wauthy G. 2001 — 'Vertition' of integumental organs in mites revisited: a case of fluctuating asymmetry — C. R. Acad. Sci., 324(5): 425-431. doi:10.1016/S0764-4469(01)01314-2 ![]()
Lindquist E.E. 1985 — External anatomy — In: Helle W., Sabelis M. W. (Eds) Spider mites – Their Biology, Natural Enemies, and Control, Amsterdam: Elsevier Science BV. pp. 3-28.
Lindquist E.E. 1998 — Evolution of phytophagy in trombidiform mites — Exp. Appl. Acarol., 22: 88-100. doi:10.1023/A:1006041609774 ![]()
Ludwig W. 1932 — Das Rechts-Links-Problem im Tierreich und beim Menschen – Mit Einem Anhang Rechts-Links-Merkmale der Pflanzen — Berlin: Springer. pp. 496
Mallik B., Kumar M.H. 1992 — Tuckerella channabasavannai sp. nov. (Acari: Tuckerellidae) from India — Int. J. Acarology, 18(2): 117-119. doi:10.1080/01647959208683941 ![]()
McDaniel B., Morihara D.K., Lewis J.K. 1975 — A new species of Tuckerella from South Dacota and a key with illustrations of all known described species — Acarologia, 17(1): 274-283.
Meyer M.K.P. (Smith), Ueckermann E.A. 1997 — A review of some species of the families Allochaetophoridae, Linotetranidae and Tuckerellidae (Acari: Tetranychoidea) — Int. J. Acarology, 23(2): 67-92. doi:10.1080/01647959708683103 ![]()
Miller L.W. 1964 — A new species of Tuckerella (Acarina, Tetranychoidea, Tuckerellidae) from Tasmania — Pap. proc. R. Soc. Tasmania, 98: 79-84.
Ochoa R. 1989 — The genus Tuckerella in Costa Rica (Acari: Tuckerellidae) — Int. J. Acarology, 15(4): 205-207. doi:10.1080/01647958908683850 ![]()
Oudemans A.C. 1937 — Band C. Tarsonemini, Stomatostigmata & Eleutherengona — In: Kritisch historisch Overzicht der Acarologie. Derde gedeelte 1805-1850., Leiden: E. J. Brill. pp. i-xxviii, 799-1348.
Perkovsky E.E., Zosimovich V.Y., Vlaskin A.P. 2010 — Rovno amber — In: Biodiversity of fossils in amber from the major world deposits, Manchester: Siri Scientifc Press. pp. 116-136.
Prasad V. 1973 — A new species of Tuckerella (Acarina: Tuckerellidae) from India — Acarologia, 15(2): 237-239.
Quiros-Gonzalez M.J., Baker E.W. 1984 — Systematics and taxonomy of Acari. 5.5 Idiosomal and leg chaetotaxy in the Tuckerellidae Baker & Pritchard; ontogeny and nomenclature — In: Griffiths D. A., Bowman C. E. (Eds) Acarology 6. Vol. 1., Chichester: Ellis Horwood Ltd. pp. 166-173.
Reuter E. 1909 — Zur Morphologie und Ontogenie der Acariden mit besonderer Berücksichtigung von Pediculopsis graminum — Acta Societatis Scientiarum Fennicae, 36(4): 1-288.
Rossi de Simons N.H. 1972 — Una nueva especie de Tuckerellidae, Tuckerella elegans sp.n. (Acari, Tetranychoidea) de la República Argentina — Rev. Inv. Agropec., 9(1): 15-22.
Schmidt A.R., Jancke S., Lindquist E.E., Ragazzi E., Roghi G., Nascimbene P.C., Schmidt K., Wappler T., Grimaldi D.A. 2012 — Arthropods in amber from the Triassic Period — Proc. Natl. Acad. Sci. USA, 109(37): 14796-14801. doi:10.1073/pnas.1208464109 ![]()
Servín R., Otero S.G. 1989 — Una nueva especie de Tuckerella de Baja California Sur, Mexico — Southwest. Entomol., 14(3): 205-212.
Sidorchuk E. 2013 — New Technique for preparation of small-sized amber samples with application to mites — In: Azar D., Engel M. S., Jarzembowski E., Krogmann L., Nel A., Santiago-Blay J. A. (Eds) Insect Evolution in an Amberiferous and Stone Alphabet. Proceedings of the 6th International Congress on Fossil Insects, Arthropods and Amber, Leiden-Boston: Brill. pp. 189-201. doi:10.1163/9789004210714_014 ![]()
Sidorchuk E.A., Klimov P.B. 2011 — Redescription of the mite Glaesacarus rhombeus (Koch & Berendt, 1854) from Baltic amber (Upper Eocene): evidence for female-controlled mating — J. Syst. Palaeontol., 9(2): 183-196. doi:10.1080/14772019.2011.566585 ![]()
Sidorchuk E.A., Norton R.A. 2010 — Redescription of the fossil oribatid mite Scutoribates perornatus, with implications for systematics of Unduloribatidae (Acari: Oribatida) — Zootaxa, 2666: 45-67.
Sidorchuk E.A., Norton R.A. 2011a — The fossil mite family Archaeorchestidae (Acari, Oribatida) I: redescription of Strieremaeus illibatus and synonymy of Strieremaeus with Archaeorchestes — Zootaxa, 2993: 34-58.
Sidorchuk E.A., Norton R.A. 2011b — The fossil mite family Archaeorchestidae (Acari, Oribatida) II: redescription of Plategeocranus sulcatus and family-group relationships — Zootaxa, 3051: 14-40.
Sidorchuk E.A., Schmidt A.R., Ragazzi E., Roghi G., Lindquist E.E. 2014 — Plant-feeding mite diversity in Triassic amber (Acari: Tetrapodili) — J. Syst. Palaeontol., 13(2): 129-151. doi:10.1080/14772019.2013.867373 ![]()
Sidorchuk E.A., Vorontsov D.D. 2014 — Computer-aided drawing system—substitute for camera lucida — Acarologia, 54(2): 229-239.
Sidorchuk E.A., Vorontsov D.D. — Microscopic preparation of amber [Internet] — [23 February 2016]; [March 10, 2017] Available from: http://www.amber.openscience.ru/index.php?page=overview&lang=en ![]()
Southcott R.V., Lange R.T. 1971 — Acarine and other microfossils from the Maslin Eocene, South Australia — Rec. S. Aust. Mus., 16(7): 1-21.
Travé J., Vachon M. 1975 — François Grandjean 1882-1975 (Notice biographique et bibliographique) — Acarologia, 17(1): 1-19.
Tucker R.W.E. 1926 — Some South African mites, mainly Tetranychidae and Eriophyidae — Cape Town: Division of Entomology. pp. 15
Van Valen L. 1962 — A study of fluctuating asymmetry — Evolution, 16(2): 125-142.
Walter D.E., Lindquist E.E., Smith I.M., Cook D.R., Krantz G.W. 2009 — Order Trombidiformes — In: Krantz G.W., Walter D.E. (Eds) A Manual of Acarology, Lubbock, Texas: Texas Tech University Press. pp. 233-420.
Wauthy G., Bauchau V., Backeljau T. 1991 — What are vertitions? — In: Dusbabek F., Bukva V. (Eds) Modern Acarology, Prague: Academia. pp. 311-318.
Wauthy G., Noti M.I., Leponce M., Bauchau V. 1999 — Taxy and variations of leg setae and solenidia in Tetranychus urticae (Acari, Prostigmata) — Acarologia, 39(3): 233-255.
Weitschat W., Wichard W. 2002 — Atlas of plants and animals in Baltic amber — Munich: Verlag Dr Friedrich Pfeil. pp. 256
Womersley H. 1940 — Studies in Australian Acarina Tetranychidae and Trichadenidae — Trans. R. Soc. S. Aust., 64(2): 233-265.
Womersley H. 1957 — A new species of Tuckerella (Acarina, Tetranychoidea, Tuckerellidae) from South Australia — Trans. R. Soc. S. Aust., 80: 73-75.



2017-04-19
Date accepted:
2017-08-10
Date published:
2018-01-26
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Sidorchuk, Ekaterina A. and Khaustov, Alexander A.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)