



The first record of the genus Myrmozercon Berlese (Acari: Mesostigmata: Laelapidae) in the Neotropical region and a description of a new species
Trach, Viacheslav A.1 and Khaustov, Alexander A.2
1✉ Department of Zoology, I. I. Mechnikov Odessa National University, Shampanskij Al. 2, Odessa, 65058, Ukraine.
2Tyumen State University, Semakova Str. 10, Tyumen, 625003, Russia.
2018 - Volume: 58 Issue: 1 pages: 41-51
https://doi.org/10.24349/acarologia/20184226ZooBank LSID: C4E25B6B-4B6D-4C25-9DA8-2B457C74A8BE
Keywords
Abstract
The family Laelapidae comprises a multitude of morphologically and behaviourally diverse mites that are free living or associated with arthropods (including ants), mammals, or birds (Lindquist et al., 2009). Worldwide, this family comprises about 90 genera and 1300 species (Beaulieu et al., 2011).
Many laelapid species of the genera Cosmolaelaps Berlese, 1903, Gaeolaelaps Evans and Till, 1966, Gymnolaelaps Berlese, 1916, Holostaspis Kolenati, 1858, Laelaspis Berlese, 1903, Myrmozercon Berlese, 1902 and Reticulolaelaps Costa, 1968 were described from ants or their nests (Bregetova 1977; Joharchi et al. 2012; Joharchi and Halliday 2013; Joharchi and Moradi 2013; Joharchi and Babaeian 2015).
Berlese (1902) erected the genus Myrmozercon and designated M. brevipes as its type species. Later on Berlese (1903) described genus Myrmonyssus with three new species and created subgenus Laelaspulus in the genus Myrmonyssus (Berlese, 1904). Rosario and Hunter (1988) synonymised Myrmonyssus to Myrmozercon, and later Shaw and Seeman (2009) synonymised the subgenus Laelaspulus and monotypic genus Parabisternalis Ueckermann and Loots, 1995 to Myrmozercon.
Myrmozercon currently includes 29 extant species and one fossil unnamed species from Eocene Baltic amber (Dunlop et al. 2014; Joharchi et al. 2015, 2017). The extant species are known from the Palaearctic (15 species), Australasia (5 species), Afrotropic (4 species) and Nearctic (4 species), but so far are unknown from Indomalaya and Neotropic regions (Joharchi and Moradi 2013; Joharchi et al. 2015, 2017; Kontschán and Seeman 2015). Myrmozercon species are associated with ants of the genera Camponotus Mayr, 1861 (Michael 1891; Berlese, 1903; Rosario and Hunter 1988; Joharchi et al. 2011); Crematogaster Lund, 1831 (Vitzthum 1939; Sellnick 1941; Hunter and Hunter 1963; Trach and Khaustov 2011; Joharchi et al. 2017); Formica Linnaeus, 1758 (Babaeian et al. 2013); Iridomyrmex Mayr, 1862 (Banks 1916; Hull 1923); Messor Forel, 1890 (Berlese 1903; Babaeian et al. 2014; Joharchi and Moradi 2013); Monomorium Mayr, 1855 (Ghafarian et al. 2013); Myrmica Latreille, 1804 (Joharchi et al. 2015); Myrmecocystus Wesmael, 1838 (Karawajew 1909); Pheidole Westwood, 1839 (Joharchi and Moradi 2013); Polyrhachis Smith F., 1857 (Walter 2003; Shaw and Seeman 2009); Tapinoma Förster, 1850 (Berlese 1902; Karawajew 1909; Kontschán and Seeman 2015).
During the study of arthropod-associated mites in the Chilean part of Patagonia, a new species of the genus Myrmozercon was revealed. The aim of this article is to describe this new species and to provide a key to non-Palaearctic species of Myrmozercon.
Ants were sampled by suction and transferred into vials containing 96% ethanol. Mites from alcohol sediments were extracted and mounted in Hoyer’s medium. Morphology of mites was studied with the aid of a compound microscope, a Mikmed-1 Lomo equipped with a binocular head AU–12, ocular micrometer AM9–2 and digital camera DCM900. The morphological nomenclature generally follows Evans and Till (1965). All pore-like structures, glandular openings (solenostomes) and poroids (lyrifissures) are named as pores. Dorsal setae were labelled according to the system of Lindquist and Evans (1965), although we stress that our designations are preliminary. Palpal and leg chaetotaxy follows Evans (1963a, b, 1969). Measurements are given in micrometres (μm) for the holotype and paratypes (in parentheses, minimum to maximum). Lengths of shields were measured from the anterior to posterior shield margins along the midline. The length of the second cheliceral segment was measured from the base to the apex of the fixed digit. Length of legs was taken from the base of the coxa to the apex of the tarsus, excluding the ambulacrum.
The holotype is deposited in the collections of the Museum of Zoology, I. I. Mechnikov Odessa National University; paratypes are deposited in the Museo de Zoología Universidad de Concepción (Concepción, Chile), the Department of Zoology I. I. Mechnikov Odessa National University (Odessa, Ukraine), the Tyumen State University Museum of Zoology (Tyumen, Russia).
Myrmozercon Berlese, 1902: 699. Type species Myrmozercon brevipes Berlese, 1902, by monotypy.Myrmonyssus Berlese, 1903: 16. Type species Myrmonyssus diplogenius Berlese, 1903, designated by Berlese, 1904 (synonymy by Rosario and Hunter, 1988).Myrmonyssus (Laelaspulus) Berlese, 1904: 437. Type species Myrmozercon acuminatus Berlese, 1903, by original designation (synonymy by Shaw and Seeman, 2009).Parabisternalis Ueckermann and Loots, 1995: 35. Type species Parabisternalis yemeni Ueckermann and Loots, 1995, by original designation (synonymy by Shaw and Seeman, 2009).
Diagnosis — The diagnosis of genus Myrmozercon follows that of Shaw and Seeman (2009) with modifications of Joharchi and Moradi (2013), and Kontschán and Seeman (2015).
ZOOBANK: 1B9CCEB5-C623-4071-9781-1C7CD68CE780 ![]()
Diagnosis — Dorsal shield broadly oval, not covering entire idiosoma; hypotrichous; with 29–30 pairs of setae; cuticle around dorsal shield sclerotised and striated; soft cuticle on posterior margin of idiosoma smooth; dorsal cuticle not hypertrichous. Sternal shield entire; setae st1–st5 present; anal shield subtriangular, without anterolateral extensions, with circum-anal setae only; peritremes short; peritrematal shields well developed, fused with dorsal shield anteriorly; ventral soft cuticle hypertrichous. Fixed cheliceral digit reduced and edentate; pilus dentilis absent; movable cheliceral digit edentate; epistome triangular, smooth; chaetotaxy of palps (from trochanter to genu 1–4–6); deutosternum with 19–21 transverse rows of deutosternal denticles (8–25 denticles per row); palp coxal seta present, simple. Chaetotaxy of legs I–IV (from coxa to tibia): 2–6–14–13/15–13; 2–5–10–11–9/10; 2–5–7/8–11–9/10; 1–5–8–11–9/10; legs long, legs I and IV subequal, longer than legs II and III.
Description of female (n=4) — Dorsum (Figure 1A). Idiosoma oval; 921 (930 – 949) long and 809 (763–781) maximum width. Dorsal shield broadly oval; not covering entire idiosoma; 763 (763 – 791) long and 716 (707 – 735) maximum width; reticulated with distinct sigillae; hypotrichous; with 29 or 30 pairs of setae (r5 often missing) (j1, j3–j6, z1–z6, s3–s6, r3–r5, J2–J5, Z2–Z5, S2–S5) and 15 pairs of distinguishable pores; pores near base of j1 absent. Cuticle around of dorsal shield strongly sclerotised and striated; with variable number of setae (three-five pairs); posterolateral margins of cuticle curved ventrally. Soft cuticle on posterior margin of idiosoma smooth and weakly striated; with variable number of setae (three-four pairs). Almost all dorsal setae barbed in distal part; 53 – 103 (53 – 99) long.

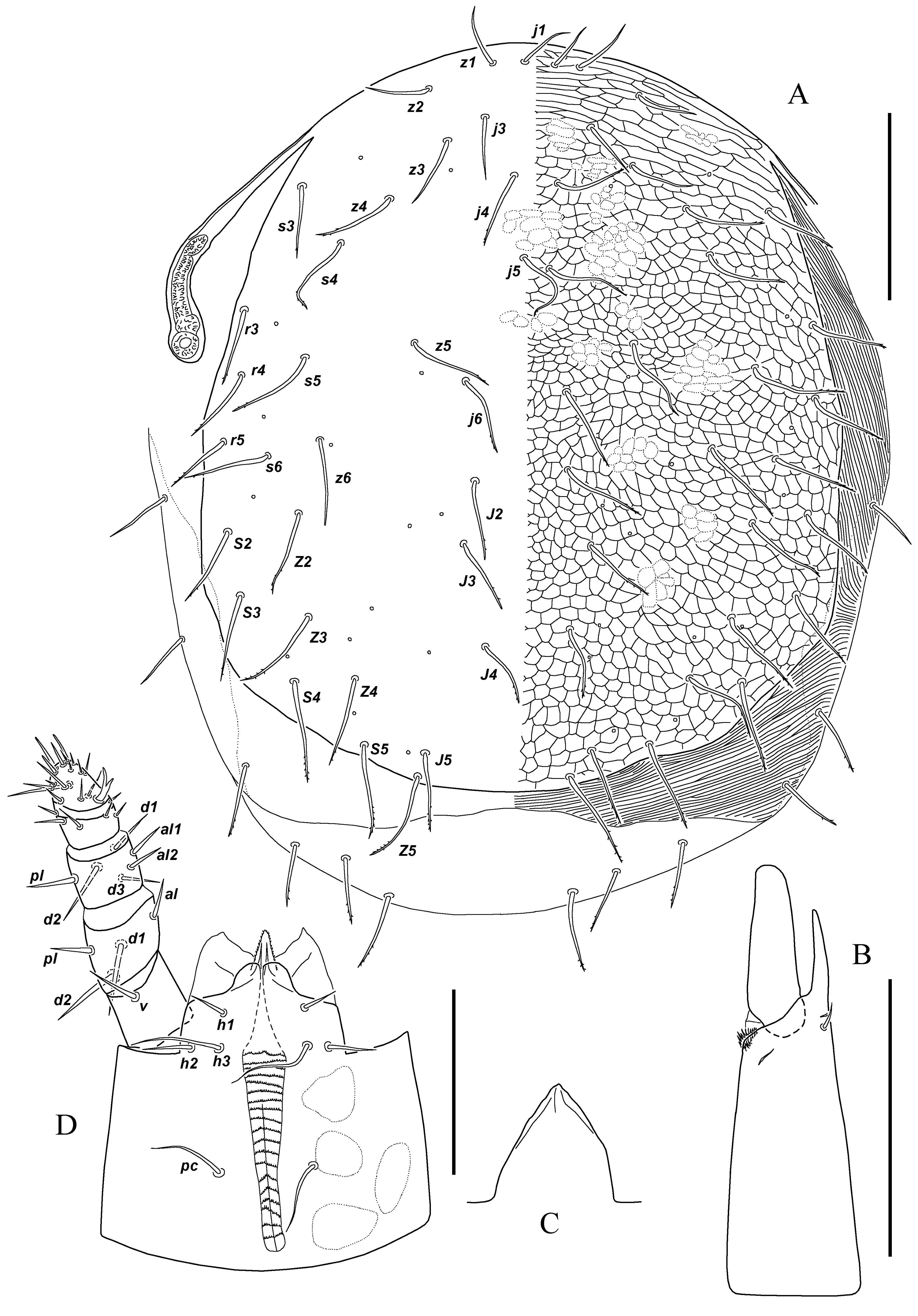
Venter (Figure 2) — Tritosternum with short, broad base and paired smooth laciniae; base of tritosternum length 19 (21 – 23), maximum width 34 (34 – 36), laciniae length 105 (103 – 113). Presternal shields fused to anterior margin of sternal shield. Sternal shield 126 (122 – 134) long midline and 176 (172 – 181) wide at narrowest level at about middle of coxae II; reticulated on anterior part and lateral margins; with three pairs of setae and two pairs of pores (iv1, iv2); pore iv1 oriented transversely, positioned posteriad seta st1, pore iv2 oriented obliquely between setae st2 and st3. Endopodal plates I–II fused with lateral margins of sternal shield alongside coxae II, endopodal III–IV connected to the latero-corner of sternal shield at anterior margin of coxae III and to parapodal platelets. Metasternal platelets absent; setae st4 and pores iv3 located on soft cuticle. Genital shield 378 (378 – 391) long and 248 (248 – 252) maximum width at level and slightly posteriorly of st5; reticulated; with setae st5. Soft cuticle at lateral margins of genital shield with one pair of pores; cuticle at posterior margin with transverse platelets (two-three pairs in different specimens). Subtriangular anal shield with smooth surface; 130 (130 – 139) long and 223 (214 – 223) maximum width; anus located in centre of shield; with circum-anal setae; pores obscure; cribrum well developed. Soft cuticle around of anal shield striated; hypertrichous; with one or two elongate irregular metapodal plates; a pair of small platelets; 22–25 pairs of setae and three-four pairs of distinguishable pores. Peritreme 139 (139 – 151) long; peritrematal shields well developed; fused to dorsal shield anteriorly; pores on peritrematal shield absent. Setae st1–st5 and about 12 pairs of setae on soft cuticle narrow and smooth; others slightly thickened and barbed (as dorsal setae); para-anal and post-anal strongly thickened and slightly barbed in distal part; measurements of setae: st1–st5 48 – 57 (44 – 57), setae on soft cuticle 44 – 80 (44 – 88), para-anal 71 (65 – 78), post-anal 84 (82 – 86).


Gnathosoma — Second cheliceral segment 139 (139 – 143) long; fixed cheliceral digit reduced and edentate; pilus dentilis absent; antiaxial and dorsal lyrifissures, cheliceral seta and arthrodial process distinct; movable cheliceral digit edentate (Figure 1B). Epistome triangular, smooth (Figure 1C). Palp 151 (147 – 155) long; numbers of setae on palp trochanter-genu: 1, 4, 6, tibia and tarsus about 23; setae aciculate and smooth; apotele two-tined, located on palp tarsus (Figure 1D). Subcapitulum 181 (181 – 193) maximum width. Deutosternum with 19–21 transverse rows of deutosternal denticles (8–25 denticles per row); marginal line of deutosternal groove weakly discernible anteriad setae h2 and h3. Corniculi short, broad, irregular shape, weakly sclerotised. Internal malae pointed and smooth, not projecting beyond tips of corniculi; labrum projecting somewhat beyond tips of internal malae. Hypostomal and palp coxal setae smooth; measurements of setae: h1 21 (21 – 23), h2 27 (27 – 29), h3 48 (44 – 46), pc 44 (40 – 44).
Legs (Figures 3A–D) — Lengths: I 1051 (1051 – 1097), II 716 (707 – 735), III 884 (865 – 893), IV 1042 (1042 – 1079). Number setae on some leg segments abnormal for Laelapidae (on femora I–IV, genua III–IV, and, sometimes, genu I and tibiae III–IV). Some leg segments with variable setation. Chaetotaxy of holotype: leg I: coxa 2, trochanter 6 (1 0/1 1/2 1), femur 14 (2 3/2 2/3 2), genu 14 (2 3/3 3/1 2), tibia 13 (2 3/2 3/1 2); leg II: coxa 2, trochanter 5 (1 0/1 0/2 1), femur 10 (2 2/1 2/2 1), genu 11 (2 3/1 2/1 2), tibia 10 (2 2/1 2/1 2), tarsus 18 (3 3/2 1/1 3/2 3); leg III: coxa 2, trochanter 5 (1 0/1 0/2 1), femur left 7 (1 2/1 2/1 0), femur right 8 (1 2/2 2/1 0), genu 11 (2 2/1 3/1 2), tibia left 10 (2 1/1 3/1 2), tibia right 9 (2 1/1 2/1 2), tarsus 18 (3 3/2 1/1 3/2 3); leg IV: coxa 1, trochanter 5 (1 0/1 0/2 1), femur 8 (1 2/1 2/2 0), genu 11 (2 2/1 3/1 2), tibia 10 (2 1/1 3/1 2), tarsus 18 (3 3/2 1/1 3/2 3). In paratypes genu I with 13 (2 3/2 3/1 2; av3 lacking), 14 (2 3/3 3/1 2) or 15 setae (2 3/3 3/2 2; added pv2); tibia II with 10 (2 2/1 2/1 2) or 9 setae (1 2/1 2/1 2; al1 lacking); femur III with 7 setae (1 2/1 2/1 0; av1 lacking); tibia with 9 (2 1/1 2/1 2) or 10 (2 1/1 3/1 2; pd3 added) setae; tibia IV with 10 (2 1/1 3/1 2) or 9 setae (2 1/1 2/1 2; pd2 lacking). All setae smooth; some thickened. Pretarsi I–IV similar in shape; claws apparently absent; ambulacral pads well-developed. Genital structures — Insemination ducts opening between bases of coxae III and IV. Tubuli sclerotised; slightly curved; 2–3 in diameter (Figure 2). Sacculus is indiscernible.


Type material — Female holotype, slide No 30–10–2015/01, Chile, Patagonia, vicinity of Puerto Natales, on Camponotus sp. collected from rotten log of Nothofagus sp., 51°56'55.2''S, 72°23'24.8''W, 30 October 2015, A.A. Khaustov coll.; paratypes: 3 females, same data.
Etymology — The name of a new species refers to its distribution in Patagonia.
Differential diagnosis — According to the presence of palpcoxal setae and setae st4, anal shield without anterolateral extensions, absence of hypotrichous leg setation and long legs, Myrmozercon patagonicus n. sp. belongs to the group of species referred to as “Myrmozercon sensu lato” by Joharchi et al. (2015).
By the hypotrichous dorsal shield, hypertrichous ventral soft cuticle and by its large size, M. patagonicus n. sp. is similar to M. antennophoroides (Berlese, 1904), M. beardae Shaw and Seeman, 2009, M. hunteri Joharchi, Babaeian and Seeman, 2015 and M. rotundiscutum Rosario and Hunter, 1988.
Myrmozercon patagonicus n. sp. differs from M. antennophoroides by the shape of its body (oval in M. patagonicus n. sp., obovoid in M. antennophoroides), shape of the genital shield (rounded posteriorly in M. patagonicus n. sp., with notch in M. antennophoroides), size and shape of the anal shield (large and subtriangular in M. patagonicus n. sp., small and indistinct in M. antennophoroides).
It differs from M. beardae by the number and shape of setae on dorsal shield (29–33 pairs of mainly barbed setae in M. patagonicus n. sp., 23–25 pairs of smooth setae in M. beardae), presence of setae st4 and st5 (absent in M. beardae), shape of setae on soft ventral cuticle (simple or barbed in M. patagonicus n. sp., simple or spinose in M. beardae), shape of the anal shield (anus located in centre of shield in M. patagonicus n. sp., in posterior part of shield in M. beardae), number setae on palps (trochanter with one seta, genu with six setae in M. patagonicus n. sp., trochanter with two setae, genu with four-five setae in M. beardae), presence of only one seta on coxa IV (two setae in M. beardae).
The new species is also very similar to M. hunteri in the number of setae on dorsal shield, shape of anal shield, thickened circum-anal setae and chaetotaxy of palps. M. patagonicus n. sp. differs from M. hunteri by hypotrichous dorsal cuticle (hypertrichous in M. hunteri), fusion of dorsal shield with peritrematal shields (free in M. hunteri), shape of circum-anal and leg setae (pointed in M. patagonicus n. sp., blunt in M. hunteri), and shape of palpcoxal setae (simple in M. patagonicus n. sp., two-tined in M. hunteri).
Myrmozercon patagonicus n. sp. is distinguished from M. rotundiscutum in having 29–30 pairs of setae on the dorsal shield (27 pairs in M. rotundiscutum), hypotrichous dorsal cuticle (hypertrichous in M. rotundiscutum), dorsal shield fused with peritrematal shields (free and weakly developed peritrematal shields in M. rotundiscutum), having 19–21 transverse rows of deutosternal denticles on deutosternum (11 in M. rotundiscutum) and six setae on palp genu (five in M. rotundiscutum).
Myrmozercon patagonicus n. sp. also differs from all congeners by the number setae on leg segments. The presence of 14 setae on femur I is also known in M. burwelli Shaw and Seeman, 2009, M. hunteri and M. rotundiscutum; the presence of 14 or 15 setae on genu I is unknown in other species. The presence of 10 setae on femur II is known in M. beardae, M. iainkayi Walter, 2003, M. iranicus Babaeian and Nemati, 2014, and M. rotundiscutum. Seven setae on femur III occur in M. cyrusi Ghafarian and Joharchi, 2013, and M. hunteri and most specimens of M. patagonicus n. sp., but presence of eight setae on femur III is unknown in other species of Myrmozercon. The new species has 11 setae on genu III (as M. burwelli and M. rotundiscutum) and 9–10 setae on tibia III (nine setae in M. beardae, M. burwelli, M. iainkayi and M. crinitus Joharchi, 2013; 10 setae in M. hunteri and M. rotundiscutum). The presence of eight setae on femur III is also known in M. brevipes Berlese, 1902 and M. iainkayi, and 11 setae on genu IV is known in M. beardae and M. burwelli. Unfortunately, leg chaetotaxy described in detail only for 16 species of Myrmozercon.
Joharchi and Moradi (2013) presented a key to 13 species of Myrmozercon occurring in the Palaearctic Region and Joharchi et al. (2015) provided a key to the 18 species of Myrmozercon that are well-described; they also briefly diagnosed the remaining species. The following key to non-Palaearctic species of the genus Myrmozercon is based on keys of Walter (2003), Shaw and Seeman (2009) and Joharchi et al. (2015) and published descriptions and illustrations.
% mise en forme de la clé de determination
1. Leg setation hypotrichous: leg I (from femur to tibia) 8–8–8, leg II 8–8–7. Palpcoxal setae sometimes absent. Setae st4 usually absent. Anal shield usually with anterolateral extensions
...... Myrmozercon sensu stricto – 2
— Leg setation holo- or hypertrichous: leg I (from femur to tibia) at least 11–11–10, leg II at least 9–10–9. Palpcoxal setae present. Setae st4 usually present. Anal shield without anterolateral extensions
...... Myrmozercon sensu lato – 6
2. Seta j1 not the longest in podonotal part of dorsal shield
...... 3
— Seta j1 the longest in podonotal part of dorsal shield
...... 5
3. Coxa IV without setae; trochanter III with 5 setae. Most dorsal setae barbed [Yemen]
...... M. yemeni (Ueckermann and Loots, 1995)
— Coxa IV with 1 seta; trochanter III with 4 setae. All dorsal setae smooth
...... 4
4. Dorsal shield with 38 pairs of setae, posterior dorsal setae spine-like. Trochanter II with 5 setae; femur IV with 7 setae; genu IV with 8 setae [USA]
...... M. clarus (Hunter and Hunter, 1963)
— Dorsal shield with 35 pairs of setae, posterior dorsal setae simple. Trochanter II with 4 setae; femur IV with 6 setae; genu IV with 7 setae [USA]
...... M. spinosus (Hunter and Hunter, 1963)
5. Opisthonotal part of dorsal shield with 5 or 6 setae longer than others. Body length 561 [Equatorial Guinea]
...... M. eidmanni (Sellnick, 1941)
— Opisthonotal part of dorsal shield with 3 or 4 setae longer than others. Body length 418 – 429 [Equatorial Guinea]
...... M. minor (Sellnick, 1941)
6. Body length less than 1000
...... 7
— Body length more than 1300 [East Africa]
...... M. titan (Berlese, 1916)
7. Dorsal shield highly hypertrichous, covered with dense pelage of more than 200 short setae
...... 8
— Dorsal shield setation hypotrichous to only moderately hypertrichous with a maximum of about 50 pairs of setae
...... 9
8. Dorsal shield 850 long. Marginal setae uniformly short [Australia]
...... M. aequalis (Banks, 1916)
— Dorsal shield 720 long. Setae fringing posterior margin longer [Australia]
...... M. scutellatus (Hull, 1923)
9. Dorsal shield with no more than 30 pairs of setae. Coxae I–III with 2 setae; coxa IV with 1 – 2 setae
...... 10
— Dorsal shield with 39–50 pairs of setae. Coxae I–III with 6 setae; coxa IV with 4 setae [Australia]
...... M. iainkayi Walter, 2003
10. Coxa IV with 2 setae
...... 11
— Coxa IV with 1 seta
...... 12
11. Setae st4 and st5 present. Anal shield with 5 setae [USA, imported from Mexico]
...... M. chapmani (Baker and Strandtmann, 1948)
— Setae st4 and st5 absent. Anal shield with 3 setae [Australia]
...... M. beardae Shaw and Seeman, 2009
12. Dorsal shield with 24–27 pairs of setae. Ventral soft cuticle holotrichous or hypertrichous, with 9–16 pairs of setae. Peritrematal shields free from dorsal shield. Femur III with 5–6 setae; femur IV with 6–7 setae
...... 13
— Dorsal shield with 29–30 pairs of setae. Ventral soft cuticle hypertrichous, with 22–25 pairs of setae. Peritrematal shields fused with dorsal shield. Femur III with 7–8 setae; femur IV with 8 setae [Chile]
...... M. patagonicus n. sp.
13. Dorsal shield with 24–25 pairs of setae. Dorsal soft cuticle not hypertrichous, with about 5 pairs of setae. Ventral soft cuticle not hypertrichous, with about 9–11 pairs of setae. Femur II with 11 setae; genu II with 12 setae; tibia II with 10 setae; femur III with 6 setae; tibia III with 9 setae; femur IV with 6 setae; genu IV with 11 setae; tibia IV with 10 setae [Australia]
...... M. burwelli Shaw and Seeman, 2009
— Dorsal shield with 27 pairs of setae. Dorsal soft cuticle hypertrichous, with about 15 pairs of setae. Ventral soft cuticle hypertrichous, with about 16 pairs of setae. Femur II with 9 setae; genu II with 11 setae; tibia II with 11 setae; femur III with 5 setae; tibia III with 10 setae; femur IV with 7 setae; genu IV with 9 setae; tibia IV with 8 setae [USA]
...... M. rotundiscutum Rosario and Hunter, 1988
The authors are grateful to Lucila Moreno Salas (Universidad de Concepción) for assistance in transfer a type specimen to Chile and to the two anonymous reviewers for their numerous suggestions for the improvement of this paper. This research of junior author was supported by the Ministry of Education and Science of the Russian Federation, project title “Myrmecophilous Acariform Mites (Acariformes) of the South of Western Siberia”, cooperative agreement No. 6.1933.2014/K.
Babaeian E., Joharchi O., Saboori A. 2013 — A new Myrmozercon Berlese (Acari: Mesostigmata: Laelapidae) associated with ant from Iran — Acarologia, 53(4): 453-460. doi:10.1051/acarologia/20132109 ![]()
Babaeian E., Seraj A., Nemati A. 2014 — Description of a new ant-associated species (Acari: Mesostigmata: Laelapidae) from Iran — Acarologia, 54(2): 221-228. doi:10.1051/acarologia/20142129 ![]()
Banks N. 1916 — Acarians from Australian and Tasmanian ants and ant nests — Trans. R. Soc. S. Aust., 40: 224-240.
Beaulieu F., Dowling A.P.G., Klompen H., Moraes de G.J., Walter D.E. 2011 — Superorder Parasitiformes Reuter, 1909. In: Zhang Z.Q. (Ed.). Animal Biodiversity: An outline of higher-level classification and survey of taxonomic richness — Zootaxa, 3148: 123-128.
Berlese A. 1902 — Specie di Acari nuovi — Zool. Anz., 25: 697-700.
Berlese A. 1903 — Diagnosi di alcuni nuove specie di Acari italiani, mirmecofili e liberi — Zool. Anz., 27: 12-28.
Berlese A. 1904 — Illustrazione iconografica degli Acari mirmecofili — Redia, 1: 299-474.
Bregetova N.G. 1977 — Fam. Laelaptidae Berlese, 1892 — In: Gilyarov M.S., Bregetova N.G. (Eds). A Key to the Soil-Inhabiting Mites of the Mesostigmata. Leningrad: Nauka. p. 483-554. [In Russian]
Dunlop J.A., Kontschán J., Walter D.E., Perrichot V. 2014 — An ant-associated mesostigmatid mite in Baltic amber — Biol. Lett., 10: 1-4. doi:10.1098/rsbl.2014.0531 ![]()
Evans G.O. 1963a — Observations on the chaetotaxy of the legs in the free-living Gamasina (Acari: Mesostigmata) — Bull. Br. Mus. (Nat. Hist.), Zool., 10: 277-303. doi:10.5962/bhl.part.20528 ![]()
Evans G.O. 1963b — Some observations on the chaetotaxy of the pedipalps in the Mesostigmata (Acari) — Ann. Mag. Nat. Hist. (Ser. 13), 6: 513-527. doi:10.1080/00222936308651393 ![]()
Evans G.O. 1969 — Observations on the ontogenetic development of the chaetotaxy of the tarsi of legs II–IV in the Mesostigmata (Acari) — In: Evans G.O. (Ed.). Proceedings of the 2nd International Congress of Acarology, 1967; Budapest: Akadémiai Kiadó. p. 195-200.
Evans G.O., Till W.M. 1965 — Studies on the British Dermanyssidae (Acari: Mesostigmata): Part I. External morphology — Bull. Brit. Mus. (Nat. Hist.) Zool., 13(8): 249-294. doi:10.5962/bhl.part.16752 ![]()
Ghafarian A., Joharchi O., Jalalizand A., Jalaeian M. 2013 — A new species of Myrmozercon Berlese (Acari, Mesostigmata, Laelapidae) associated with ant from Iran — Zookeys, 272: 21-28. doi:10.3897/zookeys.272.4404 ![]()
Hull J.E. 1923 — New myrmecophilous gamasids — Ann. Mag. Nat. Hist., 9: 610-617. doi:10.1080/00222932308632984 ![]()
Hunter P.E., Hunter C.A. 1963 — The genus Myrmonyssus with descriptions of two new species (Acarina: Laelaptidae) — Acarologia, 5: 335-341.
Joharchi O., Arjomandi E., Trach V.A. 2017 — A new species of Myrmozercon Berlese (Acari: Mesostigmata: Laelapidae) associated with an arboreal ant (Formicidae: Crematogaster) from Iran — Acarologia, 57: 725-730. doi:10.24349/acarologia/20174190 ![]()
Joharchi O., Babaeian E. 2015 — A new species of Reticulolaelaps Costa (Acari: Laelapidae) associated with Tapinoma sp. (Hymenoptera: Formicidae) from Iran, with a review of the world species — Acarologia, 55(1): 33-40. doi:10.1051/acarologia/20152146 ![]()
Joharchi O., Babaeian E., Seeman O. 2015 — Review of the genus Myrmozercon Berlese (Acari: Laelapidae), with description of a new species from Iran — Zootaxa, 3955(4): 549-560. doi:10.11646/zootaxa.3955.4.6 ![]()
Joharchi O., Halliday B. 2013 — A new species and new records of Gymnolaelaps Berlese from Iran (Acari: Laelapidae), with a review of the species occurring in the Western Palaearctic Region — Zootaxa, 3646(1): 39-50. doi:10.11646/zootaxa.3646.1.3 ![]()
Joharchi O., Halliday B., Saboori A. 2012 — Three new species of Laelaspis Berlese from Iran (Acari: Laelapidae), with a review of the species occurring in the Western Palaearctic Region — J. of Nat. Hist., 46: 1999-2018. doi:10.1080/00222933.2012.707240 ![]()
Joharchi O., Halliday B., Saboori A., Kamali K. 2011 — New species and new records of mites of the family Laelapidae (Acari: Mesostigmata) associated with ants in Iran — Zootaxa, 2972: 22-36.
Joharchi O., Moradi M. 2013 — Review of the genus Myrmozercon Berlese (Acari: Laelapidae), with description of two new species from Iran — Zootaxa, 3686(2): 244-254. doi:10.11646/zootaxa.3686.2.6 ![]()
Lindquist E.E., Krantz G.W., Walter D.E. 2009 — Order Mesostigmata — In: Krantz, G.W., Walter, D.E. (Eds). A Manual of Acarology, 3rd Edition. Lubbock: Texas Tech University Press. p. 124-232.
Karawajew W. 1909 — Myrmekophilen aus Transkaspien — Russkoe Entomologicheskoe Obozrenie, 9: 227-237.
Kontschán J., Seeman O.D. 2015 — Rediscovery and redescription of Myrmozercon brevipes Berlese, 1902 (Acari: Mesostigmata: Laelapidae) — Acarologia, 55: 19-31. doi:10.1051/acarologia/20152151 ![]()
Lindquist E.E., Evans G.O. 1965 — Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina (Acarina: Mesostigmata) — Mem. Ent. Soc. Can., 47: 1-64. doi:10.4039/entm9747fv ![]()
Michael A.D. 1891 — On the association of gamasids with ants — Proc. Zool. Soc. Lond., 43: 638-653.
Rosario R.M., Hunter P.E. 1988 — The genus Myrmozercon Berlese, with description of two new species (Acari: Mesostigmata: Laelapidae) — J. Parasitol., 74(3): 466-470. doi:10.2307/3282057 ![]()
Sellnick M. 1941 — Milben von Fernando Poo — Zool. Anz., 136: 221-228.
Shaw M.D., Seeman O.D. 2009 — Two new species of Myrmozercon (Acari: Laelapidae) from Australian ants (Hymenoptera: Formicidae) — Zootaxa, 2025: 43-55.
Trach V.A., Khaustov A.A. 2011 — A myrmecophilous mite Myrmozercon tauricus sp. n. of the family Laelapidae (Acari, Mesostigmata) from Ukraine — Vestn. zool., 45(1): 23-27.
Ueckermann E.A., Loots G.C. 1995 — A new laelapid genus and species (Acari: Parasitiformes: Laelapidae) from Yemen — Afr. Entomol., 3: 35-38.
Vitzthum G. 1941 — Ein Ameisengast (Acar.) — Mitt. Deut. Entomol. Gesell. Ber., 6: 89-94.
Walter D.E. 2003 — A new mite from an arboreal ant (Formicidae: Polyrhachis sp.): Myrmozercon iainkayi sp. nov. (Mesostigmata: Laelapidae) — Int. J. Acarol., 29: 81-85. doi:10.1080/01647950308684325 ![]()



2016-11-23
Date accepted:
2017-08-04
Date published:
2017-12-21
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Trach, Viacheslav A. and Khaustov, Alexander A.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)