



Age-specific functional response and predation capacity of Phytoseiulus persimilis (Phytoseiidae) on the two-spotted spider mite
Fathipour, Yaghoub1 ; Karimi, Maryam2 ; Farazmand, Azadeh3 and Talebi, Ali Asghar4
1✉ Department of Entomology, Faculty of Agriculture, Tarbiat Modares University, P. O. Box 14115-336, Tehran, Iran.
2Department of Entomology, Faculty of Agriculture, Tarbiat Modares University, P. O. Box 14115-336, Tehran, Iran.
3Department of Agricultural Zoology, Iranian Research Institute of Plant Protection, Agricultural Research Education and Extension Organization (AREEO), Tehran, Iran.
4Department of Entomology, Faculty of Agriculture, Tarbiat Modares University, P. O. Box 14115-336, Tehran, Iran.
2018 - Volume: 58 Issue: 1 pages: 31-40
https://doi.org/10.24349/acarologia/20184225ZooBank LSID: FB7F0AE7-70B5-4AE5-919D-4A54CB396F8B
Keywords
Abstract
The two-spotted spider mite, Tetranychus urticae Koch (Acari: Tetranychidae) is a cosmopolitan agricultural pest, feeding on more than 1100 plant species, of which more than 150 have economic value (Bolland et al., 1998; Migeon et al., 2011; Sedaratian et al., 2011; Khanamani et al., 2013; Maleknia et al., 2016a).
Predatory mites in the family of Phytoseiidae have gained much attention, because of their important role in biological control of mite and insect pests, hence, there are intensive studies about the distribution, biology, ecology and behavioral characteristics of mites of this family (Fathipour and Maleknia, 2016). Phytoseiulus persimilis Athias-Henriot is one of the most important phytoseiid species that has been known as a very efficient biological control agent of several species of spider mites (Helle and Sabelis, 1985; Alipour et al., 2016; Maleknia et al., 2016b). According to McMurtry et al. (2013), this species has a type I life style and feeds exclusively on web-producing spider mites. Owing to the ability of P. persimilis to control spider mites, it is being mass produced and sold by many commercial biological control companies.
The performance of a predator can be affected by several attributes, two of which are the functional response and predation rate (Holling, 1959; Skalski and Gilliam, 2001). Functional response describes the relationship between a predator’s consumption rate and prey density (Solomon, 1949; Holling; 1966). Several factors such as host plant traits (Cédola et al., 2001; Fathipour et al., 2001; Skirvin & Fenlon, 2003b; Ahn et al., 2010), temperature (Skirvin and Fenlon, 2003a; Li et al., 2007; Gorji et al., 2009; Hoque et al. 2010; Jafari et al., 2012), insecticides (Poletti et al., 2007), prey stage (Hoque et al., 2010; Farazmand et al., 2012), experimental unit (Madadi et al., 2011), age of a natural enemy (Asadi et al., 2006; Li et al., 2007; Nikbin et al., 2014; Fathipour et al., 2017) and generation of a natural enemy (Khanamani et al., 2017) can affect functional response and its parameters.
There are many studies about functional response of P. persimilis (Takafuji and Chant, 1976; Everson, 1980; Fernando and Hassell, 1980; Ryoo, 1986; Sabelis and Van der Meer, 1986; Skirvin and Fenlon, 2003a; Hoque et al. 2010; Xiao and Fadamiro, 2010 ; Seiedy et al., 2012), but none of them have focused on its age-specific functional response and predation rate. Therefore, this study aimed to reveal the searching efficiency and predation capacity of P. persimilis during its whole life cycle and at different life stages. Knowledge on lifetime predation capacity of a predator can help to promote its usage in biocontrol programs. On the other hand, distinguishing which stage of P. persmilis has the maximum attack rate and the lowest handling time can help to releasing programs.
The specimens of T. urticae used in this study were obtained from the College of Agriculture, Tehran University, Iran, and reared on cucumber plants (Cucumis sativus cv. ‘Beth Alpha’) in a growth chamber at 25±1ºC, 60±5% RH and a photoperiod of 16 L:8D h.
A colony of P. persimilis was obtained from Koppert Biological System (Spidex®). This predator was reared in an arena consisting of a plastic sheet put on water-saturated foam in a 26×18×10 cm plastic box that was half-filled with water. The edges of the plastic sheet were covered with moist tissue paper to prevent predators from escaping (Walzer and Schausberger, 1999). Cucumber leaves infested with T. urticae were added to the arena three times per week.
In order to obtain predators of the same age, approximately 70-80 gravid female P. persimilis were randomly selected from the stock colony and transferred to a new arena before starting the experiments. The arena consisted of a piece of cucumber leaf with abundant T. urticae placed upside down on agar inside a 19 cm diameter Petri dish. The predatory mites were allowed to lay eggs for 12 hours, then gravid females were removed from the leaf discs. The leaf discs with the P. persimilis eggs were kept in a growth chamber at 25±1ºC, 60±5% RH and a photoperiod of 16L:8 D hours. The experimental units were monitored until the eggs hatched. Protonymphs, deutonymphs, and different ages of adult females were used for the experiments.
The experimental unit was a 3.5 cm diameter cucumber leaf disc that was placed upside down on a thin layer of 2% agar in a 6 cm diameter × 1 cm height Petri dish. Seven densities (2, 4, 8, 16, 32, 64 and 128) of T. urticae eggs were used in functional response experiments. In order to make experimental units similar to natural conditions (i.e., presence of web and arrangement of eggs), considering the average T. urticae oviposition, for the above mentioned densities of T. urticae, 1, 2, 3, 6, 12, 22 and 44 gravid females of T. urticae, respectively, were transferred onto the leaf discs with a thin paintbrush, and allowed to oviposit for 24 hours and then were removed. The deposited eggs were counted, and in cases of low or high number of eggs in each density, eggs were added or removed. The predatory mites from the same-aged colony in ages 4 (protonymph), 5 (deutonymph), 6, 10, 15, 20, 25, 30, 35 and 40 days were transferred to the leaf discs with a thin paintbrush. The sex of nymphal stages was not clear, therefore mixed sexes were considered but for the adults, only females were used in the experiments.
To prevent escape of predatory mites, the 3.5 cm diameter Petri dish was sealed with parafilm. Each treatment was replicated 10 times. After 24 h the number of intact eggs remaining was counted.
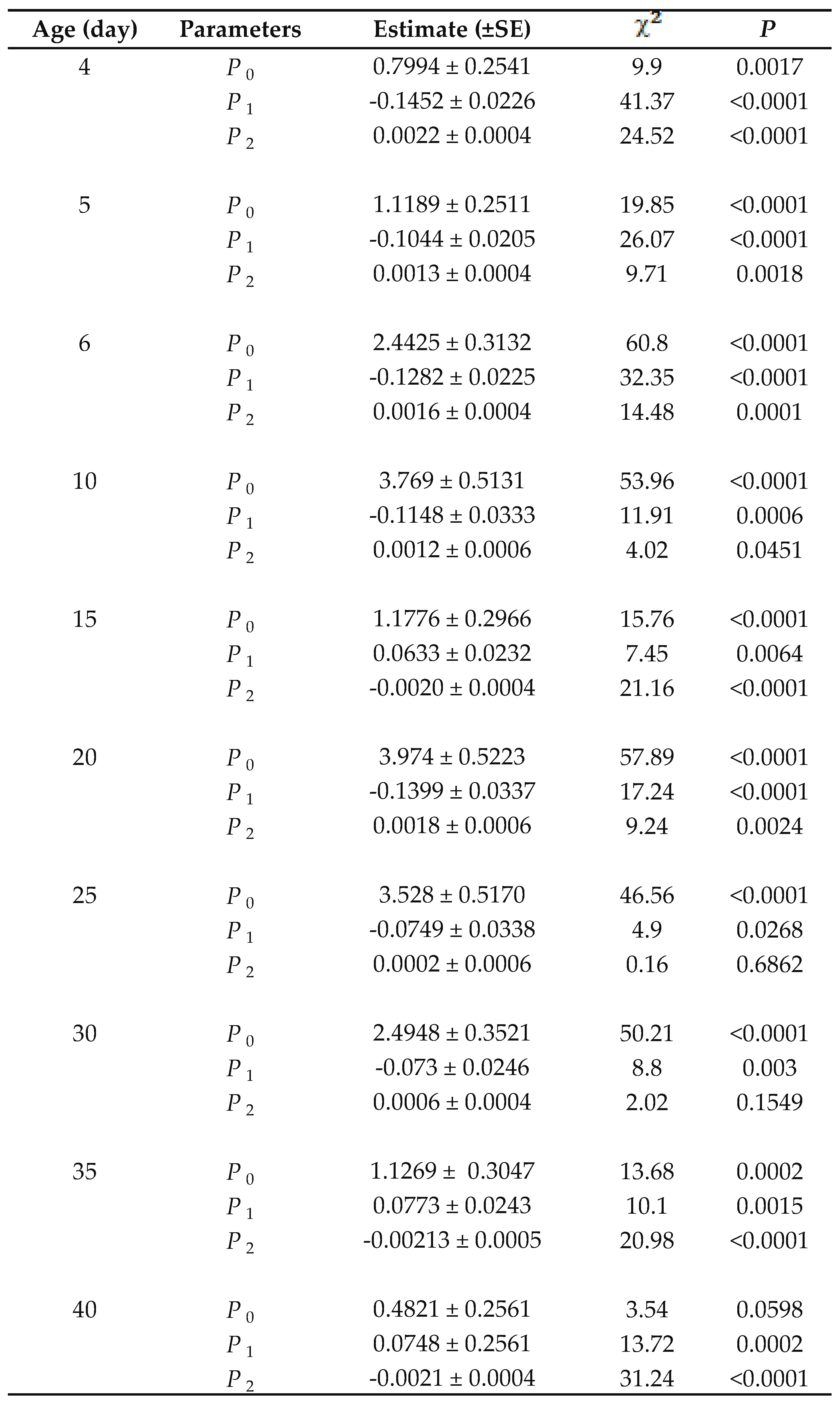
The data on functional response were analyzed in two steps (Juliano, 2001). First, the type of functional response was determined by a logistic regression of the proportion of prey consumed (Na/N 0) as a function of prey density (N0):
(1) Na / N0 = [exp(P0 + P1N0 + P2N02 + P3N03)] / [1 + exp(P0 + P1N0 + P2N02 + P3N 03)]
where P0, P1, P2 and P3 are the intercept, linear, quadratic and cubic coefficients, respectively, estimated using the method of maximum likelihood. The sign of the P1 from equation (1) can be used to distinguish the type of the functional response curve. If P1<0, it describes a type II functional response. If P1>0 and P 2<0, it shows a type III functional response (Juliano, 2001). After determining the type of functional response, the next step is to estimate the handling time and attack rate coefficients. In this study, we used an explicit deterministic model for type II functional response (Royama, 1971; Rogers, 1972):
(2) Na = Nt [1-exp(a(ThNa - T))]
where Na is the number of prey killed, Nt is the initial number of prey, T is total time available for the predator, a is attack rate and Th is the handling time. For a type III response, the attack rate is assumed to increase with host density according to the equation a = (d + bN0) / (1 + cN0) (Hassell et al., 1977). In cases where both d and c are not significantly different from 0, this leads to a = bN0 which can be inserted into equation (2).
Then, for each host density the attack coefficient (a) can be determined as a = bN0. In type III functional response, attack rate is function of prey density (Nt) and was calculated as a' = bNt. An iterative nonlinear least-squares regression was used to fit the random predator equation to data so as to estimate the parameters describing the type II response. Likewise, a non-linear least squares regression procedure was used to fit the related equation to data producing a type III response.
In order to compare the effect of different densities of the prey and different ages of predator on predation rate, analysis of variance (one-way ANOVA) was used for significant differences and Tukey’s test was used for means comparisons.
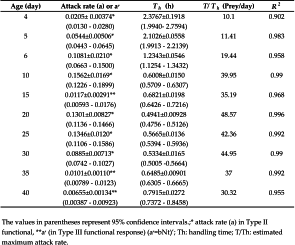
The functional response of P. persimilis of different ages at different densities of T. urticae eggs is illustrated in Fig. 1. Comparison of functional response curves revealed profound differences between predators of different ages. The results of logistic regression analyses showed that at the age of 15, 35 and 40days, the predation rate of T. urticae eggs increased, resulting in a type III functional response. The linear coefficient of equation (1) at other ages of the predator was negative and significantly different from 0 ( P<0.01), indicating a type II functional response (Table 1). The attack rate and handling time of different ages of the predatory mite are presented in Table 2. Attack rate increased from day 4 (protonymph) up to day 10 then decreased until the last day. This parameter in type III functional response is different at each density. The maximum number of T. urticae eggs consumed by P. persimilis over a 24-h period (T/Th) was 2.02 prey per hour or 48.57 prey per day at the age of 20 day. The shortest handling time (0.49 h) was also recorded at the age of 20 days and the longest handling time was estimated to be 2.38 h for protonymphs (Table 2). Handling time, showed a significant relationship with age of P. persimilis (Preg = 0.009, R2 = 0.903, Fig. 2), There was a non-linear relationship (quadratic regression) between age of P. persimilis and handling time. So that this parameter decreased of age 4 (protonymph) up to 30th (middle age of adult) then gradually increased. Hence, at younger age (protonymph) up to middle age of adult (30 days), the slope of the regression curve was negative and thereafter with increasing age, it became positive (Fig. 2).










The number of T. urticae eggs consumed by P. persimilis at different ages and varying prey densities are displayed in Table 3. The maximum prey consumption occurred at the highest density 128 of prey for all ages and was significantly higher than other densities except at the ages of 4, 6, and 35 days when feeding was not different between the densities of 64 and 128 eggs.



Generally, at low prey densities (2 and 4 eggs), prey consumption at most ages of the predator was not significantly different. With increasing prey density (16, 32, 64 and 128 eggs), the effect of age on prey consumption was more evident. Nymphal stages (age 4 and 5 days) consumed less prey than older mites. At the beginning of maturity (6 days), prey consumption at all densities became greater with increasing age. Thereafter, mean number of prey consumption indicated fluctuation in different ages and were not significantly different in many cases. Finally, it was decreased on the 40th day of predatory mite age in all densities. The highest mean number of prey consumed, at the density of 128 eggs and at the age of 20 days, was counted to be 41.7 eggs/day (Table 3) and the lowest prey consumption was 0.9 eggs/day at the density of 2 eggs and day 40. With a density of 128 prey, the lowest prey consumption was recorded within the first three days of the experiment. Also, maximum and minimum prey consumption was 46 and 7 eggs/day recorded on the 20th and 4th days of the predatory mite age, respectively.
Our findings revealed P. persimilis can show both type II and type III functional responses depending on the age of the predator supporting the hypothesis that a single predator or parasitoid species is able to exhibit different responses depending on its age. In a previous study, A. swirskii showed type II and III functional response at different ages (Fathipour et al., 2017). Furthermore, Nikbin et al. (2014) and Li et al. (2007) obtained similar results for Trichogramma brassicae Bezdenko and Scolothrips takahashii Priesner, respectively.
Some researchers reported type II functional response for phytoseiids (Cedola et al., 2001; Gotoh et al., 2004; Xiao and Fadamiro, 2010; Ahn et al., 2010; Gorji et al., 2009; Farazmand et al., 2012). In this study, immature stages of P. persimilis (protonymphs and deutonymphs) had a type II functional response to the density of prey eggs. This was in line with the findings of Fernando & Hassell (1980) for protonymphs and deutonymphs of P. persimilis. Adult females (aged 6, 10, 20, 25 and 30 days) also exhibited type II functional response. This is in agreement with Fernando & Hassell (1980) for 6-day-old adult females of P. persimilis and Xiao and Fadamiro (2010) for 2-9 days old adult females of this phytoseiid mite feeding on Panonychus citri (McGregor) nymphs. The type III functional response of 10-day-old of P. persimilis can be attributed to predators having greater reproductive rates at this time interval, hence, to compensate losing energy, females consumed a significantly higher number of prey. In a study, Moghadasi et al. (2013) found type III functional response for gravid P. persimilis females aged 3 days on T. urticae eggs.
In this study, the type III functional response in ages of 15, 35 and 40 days of the predatory mite has been observed. However, a reason for showing type III functional response in ages of 30 and 35 days might be due to more experience of females in searching for patches with higher prey densities and oviposition strategy. In another study, Fathipour et al. (2017) found type III functional response for females of Amblyseius swirskii Athias-Henriot aged 12 days.
In our study, 20-days-old females of P. persimilis showed the highest estimated maximum attack rate (T/Th) (48.57) and the lowest handling time (0.49h) compared with other ages of the predatory mite and consumption rate of P. persimilis increased with increasing prey density. This is in line with the work of Xiao and Fadamiro (2010) who showed that P. persimilis consumed higher numbers of prey in particular at high prey densities and findings of Fries and Gilstrap (1982) and Hoque et al. (2010) who showed that prey consumption by P. persimilis increased significantly with increasing densities of immature prey stages. In addition, prey consumption increased from the protonymph stage up to middle age of adult females (20 day). The same trend was observed from the protonymph up to adult female stages of P. persimilis fed with Tetranychus pacificus (Eveleigh and Chant 1981) and Typhlodromus pyri Scheuten (Sengonca et al. 2003). Likewise, previous work on A. swirskii has shown an increasing in predation rate on T. urticae at the middle-age of its life and at higher prey densities (Fathipour et al., 2017). The higher proportion of prey consumption at days fifteen, twenty, twenty-five, thirty, thirty-five and forty indicates that the predator would be more effective at controlling the T. urticae population at middle and late ages. On the other hand, the handling time showed a decreasing trend with increasing in predator age up to 30th (middle age of adult), this parameter increased again in older ages. Therefore, adult individuals at the middle and late ages are much more efficient than younger stages. This result suggests that releases of this predator may be most effective if populations can be reared and released at middle and late-stage individuals. Such releases would facilitate rapid killing of two–spotted spider mites immediately after release.
The support of this research by the Department of Entomology, Tarbiat Modares University, is greatly appreciated.
Ahn J.J., Kim, K.W., Lee J.H. 2010 — Functional response of Neoseiulus californicus (Acari: Phytoseiidae) to Tetranychus urticae (Acari: Tetranychidae) on strawberry leaves — J. Appl. Entomol., 134: 98-104. doi:10.1111/j.1439-0418.2009.01440.x ![]()
Alipour Z., Fathipour Y., Farazmand A. 2016 — Age-stage predation capacity of Phytoseiulus persimilis and Amblyseius swirskii (Acari: Phytoseiidae) on susceptible and resistant rose cultivars — Int. J. Acarol., 42(4): 224-228. doi:10.1080/01647954.2016.1171797 ![]()
Asadi R., Talebi A.A., Khalghani J., Fathipour Y., Moharramipour S., Askari Siahooei M. 2012 — Age-specific functional response of Psyllaephagus zdeneki (Hymenoptera: Encyrtidae), parasitoid of Euphyllura pakistanica (Hemiptera: Psyllidae) — J. Crop Protection, 1(1): 1-15.
Bolland H.R., Gutierrez J., Flechtmann C.H.W. 1998 — World catalogue of the spider mite family (Acari: Tetranychiade) — Leiden, Brill Academic publishers., 392 p.
Cédola C.V., Sánchez N.E., Liljesthröm G.G. 2001 — Effect of tomato leaf hairiness on functional and numerical response of Neoseiulus californicus (Acari: Phytoseiidae) — Exp. Appl. Acarol., 25: 819-831. doi:10.1023/A:1020499624661 ![]()
Eveleigh E.S., Chant D. 1981 — Experimental studies on acarine predator-prey interactions: the numerical response of immature and adult predators (Acarina: Phytoseiidae) — Can. J. Zool., 59(7): 1407-1418. doi:10.1139/z81-192 ![]()
Everson P. 1980 — The relative activity and functional response of Phytoseiulus persimilis (Acarina: Phytoseiidae) and Tetranychus urticae (Acarina: Tetranychidae): the effect of temperature — Can. Entomol., 112: 17-24.
Farazmand A., Fathipour Y., Kamali K. 2012 — Functional response and mutual interference of Neoseiulus californicus and Typhlodromus bagdasarjani (Acari: Phytoseiidae) on Tetranychus urticae (Acari: Tetranychidae) — Int. J. Acarol., 38: 369-376. doi:10.1080/01647954.2012.655310 ![]()
Fathipour Y., Kamali K., Khalghani J., Abdollahi G. 2001 — Functional response of Trissolcus grandis (Hym., Scelionidae) to different egg densities of Eurygaster integriceps (Het., Scutelleridae) and effects of wheat genotypes on it — Appl. Entomol. Phytopathol., 68: 123-136.
Fathipour Y., Karimi M., Farazmand A., Talebi A.A. 2017 — Age-specific functional response and predation rate of Amblyseius swirskii (Phytoseiidae) on two-spotted spider mite — Syst. Appl. Acarol., 22(2): 159-169.
Fathipour Y., Maleknia B. 2016 — Mite Predators. In: Omkar (ed.) Ecofriendly Pest Management for Food Security — San Diego, USA, Elsevier. pp. 329-366.
Fernando M.H.J.P., Hassell M.P. 1980 — Predator-prey responses in an acarine system — Res. Popul. Ecol., 22: 301-322. doi:10.1007/BF02530853 ![]()
Friese D.D., Gilstrap F.E. 1982 — Influence of prey availability on reproduction and prey consumption of Phytoseiulus persimilis, Amblyseius californicus, and Metaseiulus occidentalis (Acarina: Phytoseiidae) — Int. J. Acarol., 8: 85-89. doi:10.1080/01647958208683283 ![]()
Gorji M.K., Fathipour Y., Kamali K. 2009 — The effect of temperature on the functional response and prey consumption of Phytoseius plumifer (Acari: Phytoseiidae) on the two-spotted spider mite — Acarina, 17: 231-237.
Gotoh T., Nozawa M., Yamaguchi K. 2004 — Prey consumption and functional response of three acarophagous species to eggs of the two-spotted spider mite in the laboratory — Appl. Entomol. Zool., 39(1): 97-105. doi:10.1303/aez.2004.97 ![]()
Hassell M.P., Lawton J.H., Beddington J.R. 1977 — Sigmoid functional response by invertebrate predators and parasitoids — J. Anim. Ecol., 46, 249-262. doi:10.2307/3959 ![]()
Helle W., Sabelis M.W. 1985 — Spider mites, their biology, natural enemies and control (Vol.1 P.458) — Amsterdam, Elsevier.
Holling C.S. 1959 — The components of predation as revealed by a study of small mammal predation of the European pine sawfly — Can. Entomol., 91: 293-320. doi:10.4039/Ent91293-5 ![]()
Holling C.S. 1966 — The functional response of invertebrate predators to prey density — Mem. Entomol. Soc. Canada, 98(s48): 5-86. doi:10.4039/entm9848fv ![]()
Hoque M.F., Islam M.W., Khalequzzaman M. 2010 — Functional response of Phytoseiulus persimilis Athias–Henriot to Tetranychus urticae Koch: effects of prey life stages and temperature — Univ. J. Zool. Rajshahi. Univ., 29(1): 1-8.
Jafari S., Fathipour Y., Faraji F. 2012 — The influence of temperature on the functional response and prey consumption of Neoseiulus barkeri (Phytoseiidae) on two-spotted spider mite — J. Entomol. Soc. Iran., 31(2): 39-52.
Juliano S.A. 2001 — Nonlinear curve fitting: predation and functional response curves. In: Scheiner, S.M. and J. Gurevitch (eds.) — Design and Analysis of Ecological Experiments. New York, Oxford University Press, 178-196 pp.
Khanamani M., Fathipour Y., Hajiqanbar H. 2013 — Population growth response of Tetranychus urticae to eggplant quality: application of female age-specific and age-stage, two-sex life tables — Int. J. Acarol., 39: 638-648. doi:10.1080/01647954.2013.861867 ![]()
Khanamani M., Fathipour Y., Talebi A.A., Mehrabadi M., 2017 — Quantitative analysis of long-term mass rearing of Neoseiulus californicus (Acari: Phytoseiidae) on almond pollen — J. Econ. Entomol., 110(4): 1442-1450 doi:10.1093/jee/tox116 ![]()
Li D.X, Tian J., Shen Z.R. 2007 — Functional response of the predator Scolothrips takahashii to hawthorn spider mite, Tetranychus viennensis: effect of age and temperature — BioControl., 52: 41-61.
Madadi H., Mohajeri Parizi E., Allahyari H., Enkegaard A. 2011 — Assessment of the biological control capability of Hippodamia variegata (Col.: Coccinellidae) using functional response experiments — J. Pest Sci., 84: 447-455. doi:10.1007/s10340-011-0387-9 ![]()
Maleknia B., Fathipour Y., Soufbaf M. 2016a — How greenhouse cucumber cultivars affect population growth and two-sex life table parameters of Tetranychus urticae (Acari: Tetranychidae) — Int. J. Acarol., 42: 70-78. doi:10.1080/01647954.2015.1118157 ![]()
Maleknia B., Fathipour Y., Soufbaf M. 2016b — Intraguild predation among three phytoseiid species, Neoseiulus barkeri, Phytoseiulus persimilis and Amblyseius swirskii — Syst. Appl. Acarol., 21: 417-426.
Migeon A., Nouguier E., Dorkeld F. 2011 — Spider mites web: a comprehensive database for the Tetranychidae — Trends in Acarology, 557-560.
McMurtry J.A., De Moraes G.J., Sourassou N.F. 2013 — Revision of the lifestyles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies — Syst. Appl. Acarol., 18: 297-320. doi:10.11158/saa.18.4.1 ![]()
Moghadasi M., Saboori A., Allahyari H., Zahedi Golpayegani A. 2013 — Functional response of Typhlodromus bagdasarjani and Phytoseiulus persimilis (Acari: Phytoseiidae) feeding on Tetranychus urticae (Acari: Tetranychidae) on rose — Plant Pest Research, 2(4): 55-65.
Nikbin R., Sahragard A., Hosseini M. 2014 — Age-specific functional response of Trichogramma brassicae (Hymenoptera: Trichogrammatidae) Parasitizing Different Egg Densities of Ephestia kuehniella (Lepidoptera: Pyralidae) — J. Agr. Sci. Tech., 16: 1205-1216.
Poletti M., Maia A.H.N., Omoto C. 2007 C — Toxicity of neonicotinoid insecticides to Neoseiulus californicus and Phytoseiulus macropilis (Acari: Phytoseiidae) and their impact on functional response to Tetranychus urticae (Acari: Tetranychidae) — Biol. Control, 40: 30-36. doi:10.1016/j.biocontrol.2006.09.001 ![]()
Royama T. 1971 — A comparative study of models for predation and parasitism — Res. Popul. Ecol., 1: 1-90. doi:10.1007/BF02511547 ![]()
Sedaratian A., Fathipour Y., Moharramipour S. 2011 — Comparative life table analysis of Tetranychus urticae (Acari: Tetranychidae) on 14 soybean genotypes — Insect Sci., 18: 541-553. doi:10.1111/j.1744-7917.2010.01379.x ![]()
Seiedy M., Saboori A., Allahyari H., Talaei–Hassanloui R., Tork M. 2012 — Functional response of Phytoseiulus persimilis (Acari: Phytoseiidae) on untreated and Beauveria bassiana- treated adults of Tetranychus urticae (Acari: Tetranychidae) — J. Insect Behav., 25: 543-553. doi:10.1007/s10905-012-9322-z ![]()
Sengonca C., Khan H.A., Blaeser P. 2003 — Prey consumption during development as well as longevity and reproduction of Typhlodromus pyri Scheuten (Acari, Phytoseiidae) at higher temperatures in the laboratory — J. Insect Sci., 76(3): 57-64. doi:10.1046/j.1439-0280.2003.03016.x ![]()
Skalski G.T., Gilliam J.F. 2001 — Functional response with predator interference: viable alternatives to the holling type II model — Ecol., 82(11): 3083-3092.
Skirvin D.J., Fenlon J.S. 2003a — The effect of temperature on the functional response of Phytoseiulus persimilis (Acari: Phytoseiidae) — Exp. Appl. Acarol., 31: 37-49. doi:10.1023/B:APPA.0000005107.97373.87 ![]()
Skirvin D.J., Fenlon J.S. 2003b — Of mites and movement: the effect of temperature and plant connectedness on the movement of Phytoseiulus persimilis — Biol. Control, 27: 242-250. doi:10.1016/S1049-9644(03)00022-7 ![]()
Solomon M.E. 1949 — The natural control of animal populations — J. Anim. Ecol., 18: 1-35. doi:10.2307/1578 ![]()
Takafuji A., Chant D.A. 1976 — Comparative studies of two species of predacious phytoseiid mites (Acarina: Phytoseiidae), with special reference to their responses to the density of their prey — Res. Popul. Ecol., 17(2): 255-310. doi:10.1007/BF02530777 ![]()
Walzer A., Schausberger P. 1999 — Predation preferences and discrimination between con-and heterospecific prey by the phytoseiid mites Phytoseiulus persimilis and Neoseiulus californicus — BioControl., 43(4): 469-478.
XiaoY., Fadamiro H.Y. 2010 — Functional responses and prey-stage preferences of three species of predacious mites (Acari: Phytoseiidae) on citrus red mite, Panonychus citri (Acari: Tetranychidae) — Biol. Control, 53: 345-352. doi:10.1016/j.biocontrol.2010.03.001 ![]()



2017-05-04
Date accepted:
2017-08-02
Date published:
2017-12-21
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Fathipour, Yaghoub; Karimi, Maryam; Farazmand, Azadeh and Talebi, Ali Asghar
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)