



A new genus and species of Tarsonemidae (Acari: Heterostigmata) associated with Aradus betulae (Heteroptera: Aradidae) from European Russia
Khaustov, Alexander A.1 and Abramov, Vladimir V.2
1✉ Tyumen State University, Tyumen, Semakova 10, 625003 Russia.
2Gagarin str. 12, Suvorov, Tula Province, 301430, Russia.
2017 - Volume: 57 Issue: 4 pages: 1079-1087
https://doi.org/10.24349/acarologia/20174220ZooBank LSID: 0D3FFF2B-08B8-4B41-8D64-493C9D37D8EA
Keywords
Abstract
The family Tarsonemidae is one of the largest groups in Heterostigmata and currently includes 42 genera and more than 600 species (Zhang et al. 2011; Lofego et al. 2016). Members of the family are phytophagous, mycophagous, parasites and parasitoids of insects, and predators preying on mite eggs (Lindquist 1986). Tarsonemid mites inhabit soils and litter, various plants, subcortical galleries of insects, etc. Many species utilize insects for phoresy (Lindquist 1986). The insect-associated Tarsonemidae of Russia are insufficiently studied. Most of them are known as associates of subcortical insects, especially bark beetles (Khaustov 1998, 2001; Magowski and Khaustov 1999, 2006; Khaustov and Magowski 2003; Khaustov et al. 2016). During the study of mites associated with insects, a new genus and species of Tarsonemidae was revealed.The aim of this article is to describe a new genus and a new species associated with the aradid bug Aradus betulae from European Russia.
Mites were collected from under hemielytra of single specimen of an aradid bug Aradus betulae (Linnaeus, 1758) and mounted in Hoyer’s medium. The terminology follows that of Lindquist (1986), except the ventral subcapitular seta is labeled su (Seeman et al. 2016). All measurements are given in micrometers (μm) for the holotype and five female paratypes (in parentheses). For leg chaetotaxy the number of solenidia is given in parentheses. DIC micrographs were taken using the Carl Zeiss Axio Imager A2 compound microscope and digital Cameras Axiocam 506 color and Hitachi KP-HD20A. The holotypes and most paratypes of the new species are deposited in the mite collection of the Tyumen State University Museum of Zoology, Tyumen, Russia; two female paratypes of the new species are deposited in the acarological collection of the Zoological Institute of RAS, St. Petersburg, Russia.
ZOOBANK: B6B538B6-D0B8-4839-B62F-8B5C807EEDAA ![]()
Type species: Lobotarsonemus betulae n. sp.
Differential diagnosis — Females of Lobotarsonemus n. gen. resemble those of Rhynchotarsonemus Beer, 1954 from the tribe Tarsonemini in having a beaklike gnathosoma and in having legs IV much shorter than leg III. They also resemble those of Suskia Lindquist, 1986 from the tribe Steneotarsonemini in having the gnathosoma strongly prolonged, beaklike and retain seta pl" on tarsus II. Females of Lobotarsonemus differ from those of both other genera in having three large lobes on anterior part of poststernal plate, by the presence of dorsodistal projections on tarsi II and III and by absence of seta v’Fe on femorogenu IV. Females of Lobotarsonemus n. gen. also resemble those of Biscutulumnemus Lofego and Feres, 2006 from the tribe Tarsonemini in having the gnathosoma strongly prolonged, strongly reduced poststernal apodeme and retain seta pl" on tarsus II. Females of Lobotarsonemus differ from Biscutulumnemus in having three large lobes on anterior part of poststernal plate, by the presence of dorsodistal projections on tarsi II and III and by absence of seta v’Fe on femorogenu IV.
Remarks — Lobotarsonemus keys to couplet 17 of Lindquist (1986) which separates Suskia and Rhynchotarsonemus on the basis of their beak-like gnathosoma. These genera are placed in different tribes – Steneotarsonemini and Tarsonemini, respectively – showing that Lindquist (1986) thought that the beak-like gnathosoma was independently derived. Lobotarsonemus is best placed in the Steneotarsonemini because of its reduced sejugal apodeme; in the Tarsonemini this apodeme is complete. Lobotarsonemus is also closer to Suskia in its retention of a spine-like pl″ on tarsus II and solenidion φ2 on leg I, and on femur I seta l′ is not enlarged, and seta v″ is not on a finger-like apophysis.
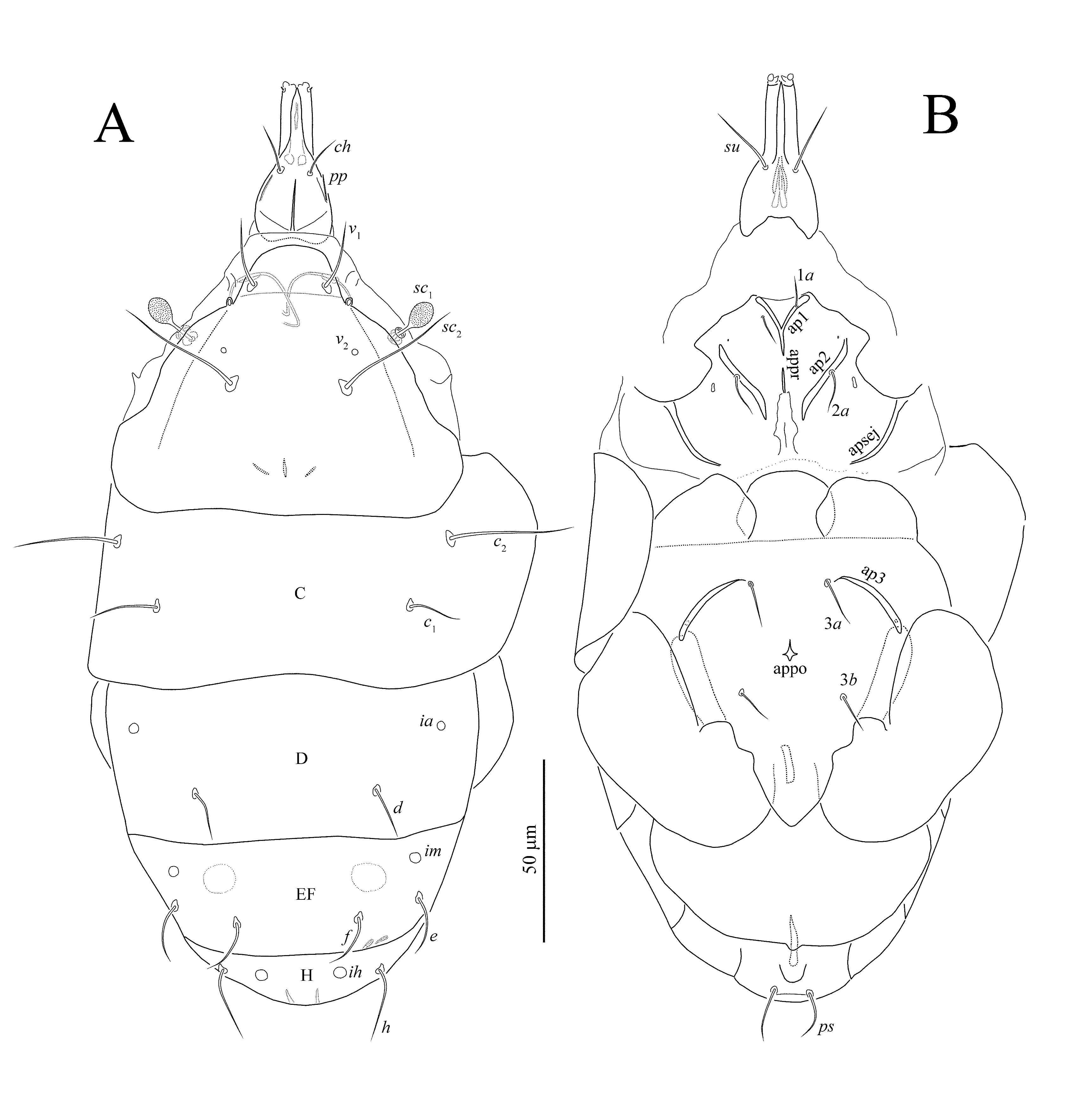
Female Gnathosoma — Capsule much longer than wide, strongly prolonged, beaklike, anteriorly; palpcoxal setae (pp) present laterally. Dorsal gnathosomal setae smooth, simple. Palpi directed anteriorly, parallel, strongly elongate, each with one minute setae and inconspicuous processes near distal extremity. Cheliceral stylets long, straight, attached to basal levers functioning in dorsoventral plane. Pharynx thin, with muscular, thinly sclerotized walls, and with conspicuous paired gland-like structures posteriorly.
Idiosoma — Dorsal shielding unornamented, with setae collectively slender, smooth. Prodorsal shield weakly extended. Stigmata projecting slightly from margins of prodorsal shield, situated closely posterolaterad setae v 1; main tracheal trunks with unsclerotized atria, and lacking sclerotized or divided postatrial structures. Bothridia present, with capitate sensilla sc 1 only basally covered by prodorsal shield. Posterior margins of tergites not conspicuously emarginate or concave. Ventral shielding with apodemes 1 forming Y-shaped juncture with prosternal apodeme. Apodemes 2 not united with prosternal apodeme. Prosternal apodeme with gap, not united with sejugal apodeme, but continuous along length to level of medial extremities of apodemes 2. Sejugal apodeme developed only laterally. Apodemes 3 not extending laterad of trochanters III, nor mediad setae 3a. Apodemes 4 absent. Poststernal apodeme strongly reduced. Anterior margin of posterior sternal plate with three large lobes (Figure 5B). Two pairs of coxal setae on metapodosomal venter; setae 3c, and 4a between legs IV, absent. Bases of legs IV moderately well-spaced, separated by interval of about three times the width of trochanter IV; tegula moderately long, rounded apically. Aggenital plate lacking setae. Pseudanal setae present.
Legs — Ambulacrum of leg I with single, typically uncinated claw. Ambulacra of legs II and III with empodium and well-developed, symmetrically paired claws. Tarsi II and III with distinct dorsodistal projections just anteriad of setae tc". Femur I without apophysis; femur II without ventral flange. Legs I and II of moderate length, with none of segments unusually elongated. Trochanter III of adult female elongate, subelliptical, plate-like, longer than femorogenu III. Leg IV elongate-cylindrical, about twice shorter than leg III, with femorogenu at least 1.5 times as long as tibiotarsus; femorogenu with one seta v′Fe, seta v′Ge absent; trochanter no longer than tibiotarsus. Number of setae and solenidia on femur, genu, tibia, and tarsus, respectively: leg I: 4-4-6(2φ) + 6(lω); leg II: 3-3-4-5(lω); leg III: 1+3-4-4. Tarsal setation of leg I lacking setae pv", pl" and (u), and with subunguinal seta spine-like; tectal longer than proral eupathidia; tarsus II with spine-like seta pl". Seta u′ on tarsi II and III spinelike, with rounded tip.
Male and larva unknown.
Species included — The genus Lobotarsonemus includes one species, L. betulae n. sp.
Distribution and habitat — Lobotarsonemus betulae n. sp. is phoretic under hemielytra of Aradus betulae (Linnaeus, 1758) in European Russia.
Etymology — The name of the new genus is a combination of two words, the Latin lobus meaning lobe, and Tarsonemus, the type genus of the family Tarsonemidae and refers to the presence of three unusual lobes on anterior margin of posterior sternal plate.
(Figures 1–5)
ZOOBANK: 37AAF394-AB56-4132-9E84-291CFC7D5826 ![]()








Description
Female (Figures 1-5) — Length of idiosoma 210 (205 – 215), width 105 (105 – 110).
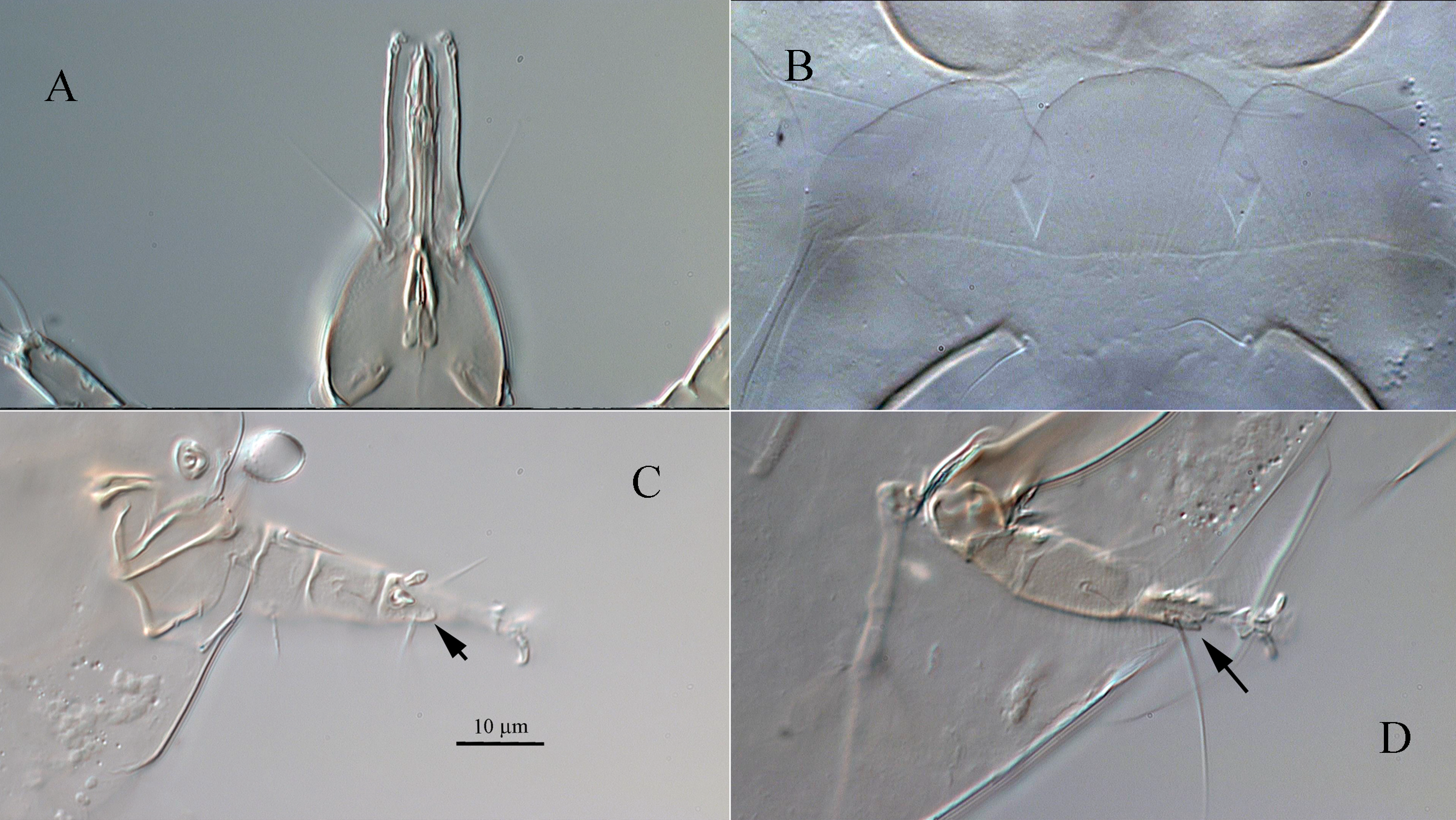
Gnathosoma (Figure 5A) — Length of gnathosomal capsule 45 (44 – 46), width 22 (20 – 23). Dorsomedian apodeme well developed, extending nearly entire distance from union basally with circumcapitular apodeme to level of insertion of dorsal gnathosomal setae (ch). Gnathosoma with dorsal pair of setae ch 14 (12 – 14) slightly shorter than subcapitular setae su 20 (18 – 20); both setae slender and smooth. Palpcoxal setae (pp) slightly shorter than ch, slender and pointed. Palps very long 23 (22 – 23).
Idiosomal dorsum (Figures 1A, 4A, C, D) — Prodorsal shield (Figure 4C) with uniform small round dimples, subtriangular, with anterior margin not hood-like over gnathosoma. Prodorsum with well-developed ω-like median apodeme. Pseudostigmatic organs capitate, ellipsoidal, finely spiculate and rounded apically. Pits v 2 located anterolaterally to bases of setae sc 2. All dorsal idiosomal setae smooth, pointed. All hysterosomal tergites with uniform small dimples (Figure 4D). Tergite EF with large, round porous areas located anteriorly to setae f. Lengths of dorsal setae: v 1 21 (20 – 21), sc 2 39 (38 – 40), c 1 18 (17 – 19), c 2 32 (31 – 33), d 15 (14 – 16), e 19 (18 – 20), f 18 (18 – 21), h 21 (19 – 22). Distances between setae: v 1–v 1 21 (21 – 22), v 2–v 2 35 (35 – 36), sc 2–sc 2 32 (31 – 32), c 1–c 1 70 (64 – 70), c 2–c 2 92 (88 – 93), c 1–c 2 32 (21 – 32), d–d 49 (44 – 50), e–e 67 (65 – 67), e–f 18 (17 – 19), f–f 33 (31 – 33), h–h 43 (41 – 44).
Idiosomal venter (Figures 1B, 4B, E, F, 5B) — All ventral plates with uniform, very small dimples. All ventral setae smooth, pointed. Pits 1b very small, in some specimens indiscernible; pits 2b large, elongate. Posterior margin of aggenital plate with short tongue-like projection. Setae 1a located just near apodemes 1 or slightly removed posteriorly; setae 2a located on apodemes 2. Lengths of ventral setae: 1a 9 (8 – 9), 2a 14 (13 – 14), 3a 14 (14 – 15), 3b 11 (11 – 12), ps 15 (14 – 15).
Legs (Figures 2, 3, 5C, D) — Lengths of legs: I 52 (50 – 53), II 53 (52 – 54), III 42 (40 – 42), IV 23 (22 – 24). Leg I (Figure 2A). Solenidia ω, φ1 and φ2 capitate; seta k 3 (3) rod-shaped, subequal to solenidion φ2. Lengths of solenidia: ω 4 (4), φ1 4 (4), φ2 3 (3). Setae d, l′ of femur, l′, v′ of genu, l′, v′ of tibia blunt-ended and smooth; other leg setae (except eupathidia) pointed and smooth. Leg II (Figures 2B, 5C). Solenidion ω 3 (3) capitate. Setae d, l′ of femur, l′, v′ of genu, and l′ of tibia blunt-ended and smooth; setae v″ of femur and v′ of tibia weakly barbed; other leg setae pointed and smooth. Leg III (Figures 3A, 5D). Setae v′ of femur and l′ of genu blunt-ended, smooth; seta u′ with basal projection; other leg setae pointed and smooth. Leg IV (Figure 3B). All leg setae pointed and smooth. Seta tc″ very long.
Type material — Female holotype, slide VA130416, Tula Province, vicinity of Suvorov town, 13 April 2016, under hemielytra of bug Aradus betulae (Linnaeus, 1758), coll. V.A. Abramov. Paratypes: 10 females, same data.
Etymology — The specific epithet of the new species is derived from the name of its phoretic host Aradus betulae.
Specimens of the newly described genus and species were collected under hemielytra of aradid bugs. In our opinion, they are phoretic rather than parasitic on insect host, because junior author collected Lobotarsonemus betulae on aradid bugs only during late April and May, also we did not find eggs, males or larvae on the insect host and not observed physogastry of females. Tarsonemid mites are known phoretic on various insects such as subcortical beetles (Coleoptera: Cerambycidae, Curculionidae), hispine beetles (Coleoptera: Chrysomelidae), fig wasps (Hymenoptera: Agaonidae), and bees (Hymenoptera: Apidae) (Lindquist 1986; Ochoa and OConnor 1996; Khaustov and Magowski 2003). The phoretic association of tarsonemid mites with Heteroptera was recorded only one time: Ceratotarsonemus teqmen Lin and Zhang, 1995 on aradid bug Mezira sp. (Lin, Zhang 1995; Ochoa et al. 1997), although members of the Coreitarsonemini are presumed to parasitic in the odiferous glands of stink bugs (Lindquist 1986). The phoresy of Lobotarsonemus betulae under hemielytra of Aradus betulae is represented the second phoretic association of Tarsonemidae with bugs of the family Aradidae and first record on bugs of the genus Aradus.
Lobotarsonemus betulae is characterized by the presence of unusual three lobes on anterior margin of poststernal plate. This character is unknown in any tarsonemid genus. Similar lobes are known only in the genus Pseudotarsonemoides Vitzthum, 1921 (subfamily Pseudotarsonemoidinae), but in Pseudotarsonemoides there are only two lobes overlapping each over, the median lobe is absent. These lobes are very likely to be independently derived as the Pseudotarsonemoidinae is a well-defined subfamily of Tarsonemidae and Lobotarsonemus is clearly a member of the Tarsoneminae.
The authors thank Dr. E.E. Lindquist (Ottawa, Canada) for valuable comments. We also thank Dr. A.C. Lofego (Universidade Estadual Paulista, São José do Rio Preto, São Paulo, Brazil) for providing information about the morphology of the genus Biscutulumnemus.
Beer R.E. 1954 — A revision of the Tarsonemidae of the Western Hemisphere (Order Acarina) — Kans. Univ.Sci. Bull., 36: 1091-1387.
Khaustov A.A. 1998 — A review of mites of the genus Pseudotarsonemoides (Acari: Heterostigmata: Tarsonemidae) of Crimea, with description of two new species — Acarina, 6(1-2): 25-30.
Khaustov A.A. 2001 — New and little known species of mites of the genus Heterotarsonemus (Acarina, Heterostigmata, Tarsonemidae) from Crimea — Vestn. Zool., 35(5): 81-84.
Khaustov A.A., Magowski W.L. 2003 — New data on tarsonemid mites (Acari: Tarsonemidae) associated with subcortical beetles (Coleoptera) in Ukraine and Russia — Acarina, 11(2): 241-245.
Khaustov A.A., Trach V.A., Bobylev A.N. 2016 — Mites (Acari) phoretic on six-toothed spruce bark beetle, Pityogenes chalcographus Linnaeus (Coleoptera: Curculionidae: Scolytinae) in Western Siberia, Russia — Acarina, 24(2): 51-65.
Lin J.-Z., Zhang Y.-X. 1995 — A new species of the genus Ceratotarsonemus DeLeon, 1956 from Fujian, China (Acari: Tarsonemidae) — Acta Zootaxon. Sinica, 20(4): 426-428.
Lindquist E.E. 1986 — The world genera of Tarsonemidae (Acari: Heterostigmata): a morphological, phylogenetic, and systematic revision, with a reclassification of family-group taxa in the Heterostigmata — Mem. Entomol. Soc. Can., 118: 1-517. doi:10.4039/entm118136fv ![]()
Lofego A.C., Feres R.J.F. 2006 — A new genus and species of tarsonemid mite (Acari: Tarsonemidae) from Brazil — Zootaxa, 1299: 45-55.
Lofego A.C., Pitton T., Resende J.M. 2016 — A new genus and new species of Tarsonemidae (Acari: Heterostig¬mata) from the Brazilian rainforests — Syst. Appl. Acarol., 21: 307-319.
Magowski W.L., Khaustov A.A. 1999 — Tarsonemus spathulaphorus, a new species of the tarsonemid mite (Acari: Tarsonemidae) from Ukraine — Internat. J. Acarol., 25(4): 243-253. doi:10.1080/01647959908684161 ![]()
Magowski W.L., Khaustov A.A. 2006 — Two new species of Tarsonemus (Acari: Tarsonemidae) associated with bark beetles (Coleoptera: Scolytidae) from Crimea, Ukraine — Zootaxa, 1359: 31-55.
Ochoa R., OConnor B.M. 1998 — A new genus of Tarsonemidae (Acari: Heterostigmata) associated with an orchid bee, Euglossa sp. (Hymenoptera: Apidae) in Brazil — Internat. J. Acarol., 22(3): 203-207. doi:10.1080/01647959608684097 ![]()
Ochoa R., Vargas C., Walter D.E., OConnor B.M. 1997 — Two new species of the genus Ceratotarsonemus (Acari: Tarsonemidae) — Internat. J. Acarol., 23(3): 177-183. doi:10.1080/01647959708683560 ![]()
Seeman O.D., Loch D.S., Knihinicki D.K., McMaugh P.E. 2016 — A new species of Steneotarsonemus (Acari: Tarsonemidae) from kikuyu grass, Pennisetum clandestinum (Poaceae), in Australia — Syst. Appl. Acarol., 21: 889-906.
Zhang Z.-Q., Fan Q.-H., Pesic V., Smit H., Bochkov A.V., Khaustov A.A., Baker A., Wohltmann A., Wen T.-H., Amrine J.W., Beron P., Lin J.-Z., Gabrys G., Husband R. 2011 — Order Trombidiformes Reuter, 1909. In: Zhang Z-Q. (ed.) Animal biodiversity: an outline of higher-level classification and survey of taxonomic richness — Zootaxa, 3148: 129-138.



2017-05-15
Date accepted:
2017-06-30
Date published:
2017-09-29
Edited by:
Auger, Philippe

This work is licensed under a Creative Commons Attribution 4.0 International License
2017 Khaustov, Alexander A. and Abramov, Vladimir V.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)