



A new species of Myrmozercon Berlese (Acari: Mesostigmata: Laelapidae) associated with arboreal ant (Formicidae: Crematogaster) from Iran
Joharchi, Omid1 ; Arjomandi, Elham2 and Trach, Viacheslav A.3
1✉ Department of Plant Protection, Yazd Branch, Islamic Azad University, Yazd, Iran.
2Department of Entomology, Faculty of Agriculture, Tarbiat Modares University, 14115–336, Tehran, Iran.
3Department of Zoology, I. I. Mechnikov Odessa National University, Shampanskij Al., 2, 65058, Odessa, Ukraine.
2017 - Volume: 57 Issue: 4 pages: 725-730
https://doi.org/10.24349/acarologia/20174190ZooBank LSID: F51400D4-BA00-47F7-B823-37D0463A30BB
Keywords
Abstract
Many species Laelapidae have been reported from ants or their nests, but the genus Myrmozercon Berlese, has a more intimate association with ants. Currently, Myrmozercon comprises 29 described species from Europe, Transcaucasia, Middle East, Central Asia, Africa, North America and Australia (Karawajew 1909; Vitzthum 1930; Hunter and Hunter 1963; Rosario and Hunter 1988; Walter 2003; Shaw and Seeman 2009; Trach and Khaustov 2011; Joharchi et al. 2011; Ghafarian et al. 2013; Joharchi and Moradi 2013; Babaeian et al. 2013, 2014; Joharchi et al. 2015). The most recent taxonomic work on the genus (Joharchi et al. 2015) revised the generic concept and its morphological attributes, provided a key to the well-described species of Myrmozercon and reviewed the poorly described species. They suggested that the most highly modified species of Myrmozercon from a monophyletic group, termed Myrmozercon sensu stricto, because it includes the type species Myrmozercon brevipes Berlese, 1902 (redescribed by Kontschan & Seeman 2015). The members of Myrmozercon sensu stricto not only share a multitude of reductive or specialized character states, but also tend to associate with ants in the genus Crematogaster (Joharchi et al. 2015).
Laelapidae associated with ants were collected in different regions of Iran over a period of eight years (2008 – 2016). Mites were removed from ants' nests by individual hand picking and by extraction from ant nesting material using Tullgren funnels. Mites were cleared in Nesbitt’s solution and mounted in Hoyer’s medium (Walter and Krantz 2009). The line drawings and examinations of the specimens were performed with a compound microscope (XS–2100 series) equipped with a drawing tube. Pencil line drawings were then scanned and traced over using Microsoft Office © Powerpoint 2003. Measurements of structures are expressed as minimum-maximum ranges in micrometres (μm). Dorsal shield length and width were taken from the anterior to posterior margins along the midline, and at its broadest point, respectively. Length and width of the sternal shield were measured from the anterior point to the posterior point at the full length and broadest point, respectively. Genital shield length and width were measured along the midline from the posterior margin of the sternal shield to the posterior margin of the genital shield, and at the level of setae st5, respectively. Leg lengths were measured from base of the coxa to the apex of the tarsus, excluding the pre-tarsus. Lengths for the fixed and movable cheliceral digits were taken from the base of the movable digit to their tips. The nomenclature used for the dorsal idiosomal chaetotaxy is that of Lindquist and Evans (1965), the leg chaetotaxy is that of Evans (1963), and names of other anatomical structures mostly follow Evans and Till (1979). We use the term "lyrifissure" to refer to slit-shaped sensilli, and "pore" for circular or oval-shaped cuticular openings of unspecified function. The holotype and one paratype of the new species are deposited in the Acarological collection, Department of Plant Protection, Yazd Branch, Islamic Azad University (YIAU). Paratypes are also deposited in the Jalal Afshar Zoological Museum, College of Agriculture, University of Tehran, Iran (JAZM) and in Museum für Naturkunde (MfN), Berlin, Germany.
Myrmozercon Berlese, 1902: 699. Type species Myrmozercon brevipes Berlese, 1902, by monotypy.Myrmonyssus Berlese, 1903: 16. Type species Myrmonyssus diplogenius Berlese, 1903, designated by Berlese, 1904 (synonymy by Rosario and Hunter 1988).Myrmonyssus (Laelaspulus) Berlese, 1904: 437. Type species Myrmozercon acuminatus Berlese, 1903, by original designation (synonymy by Shaw and Seeman 2009).Parabisternalis Ueckermann and Loots, 1995: 35. Type species Parabisternalis yemeni Ueckermann and Loots, 1995, by original designation (synonymy by Shaw and Seeman 2009).
The diagnosis of the genus Myrmozercon used here is based on Joharchi et al. (2015) and Shaw and Seeman (2009).
Myrmozercon tauricus Trach and Khaustov, 2011: 23 – Joharchi and Moradi, 2013: 250, (misidentification).
ZOOBANK: FB546BDF-8B25-4594-AD15-8CEF1D1CE9A4 ![]()
Type material — Holotype, female, Iran, Alborz province, Savojbolagh, Khoznan, 36˚71’ N, 50˚32’ E, alt. 1595 m, 15 August 2016, O. Joharchi coll., clinging to the head of Crematogaster inermis Mayr (Formicidae) (misidentification in Joharchi and Moradi, 2013) in the bark of grapevine (in YIAU). Paratypes: one female same data as holotype (in JAZM), two females, Iran, Damavand mountain, 35°52' N, 52°07' E, alt. 2422 m, 18 May 2013, O. Joharchi coll., clinging to the head of Crematogaster inermis (one in YIAU and one in MfN) (host ants are also in YIAU).
Description of female (n = 4) — Dorsal idiosoma (Figure 1A). Length 470 – 475. Dorsal shield length 440 – 449, width 316 – 325. Shield posteriorly truncate, not covering entire idiosoma, leaving a strip of striate cuticle posterior to setae J5, shield without distinct reticulate ornamentation over whole surface; with 34 pairs of setae, 20 podonotal (z2 and z6 absent), 14 opisthonotal, Z5 in soft skin posterior to shield, almost all setae slightly barbed in apical third or less, all setae on shield minute, uniform in length 12 – 15 and thickness, without unpaired or asymmetrical setae. Setae in R-series absent. Shield with 12 pairs of pore-like structures; lyrifissures near base of j1 large and slit-like.

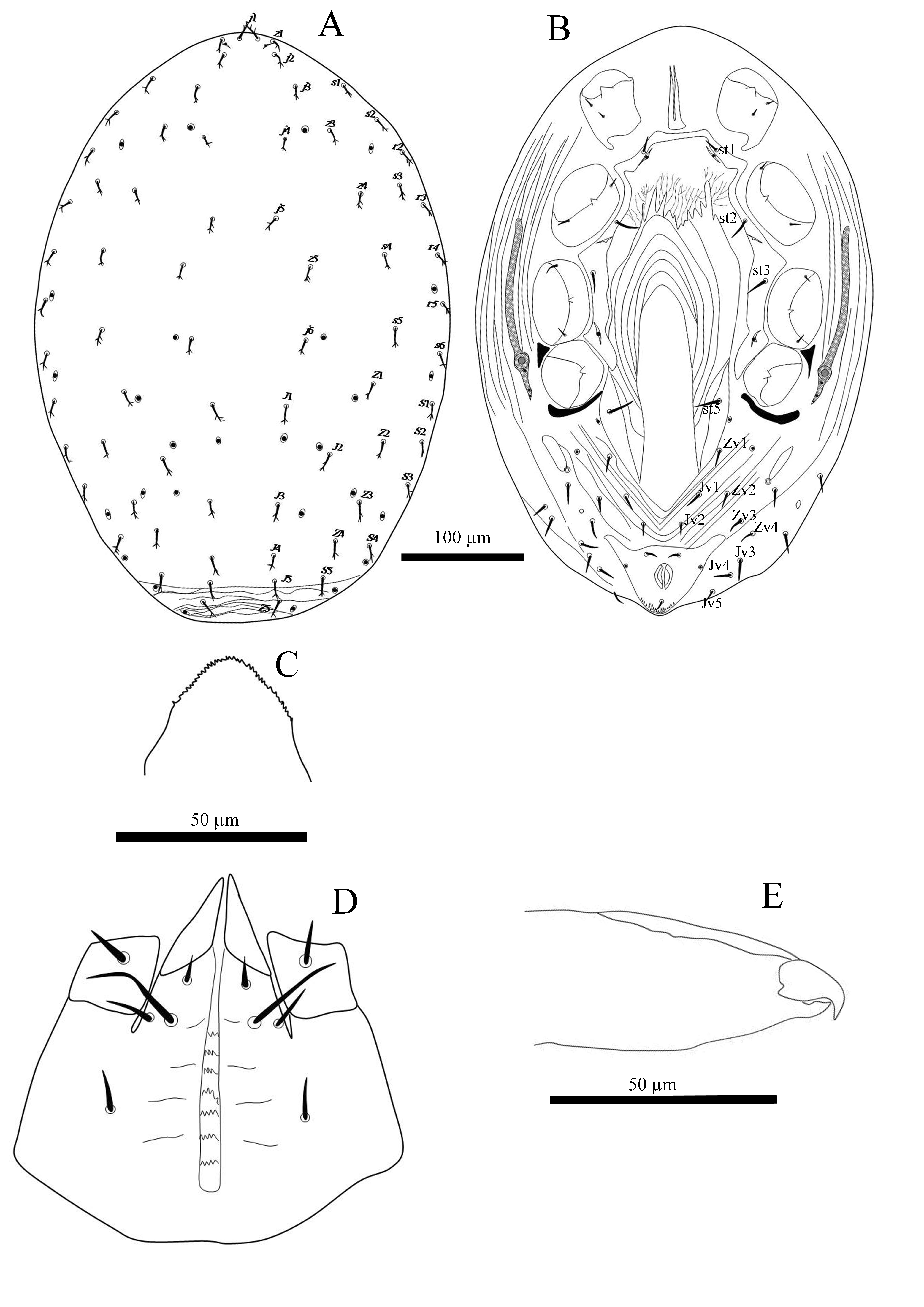
Ventral idiosoma (Figure 1B) — Tritosternum with short broad base (25 – 30 × 14 – 16 wide), laciniae 46 – 48 forked a short distance above suture, with smooth edges, strap-like and broad at base; pre-sternal shields apparently fused with sternal shield. Sternal shield (length 200 – 210) narrowest between coxae II (106 – 108) widest between coxae II and III (166 – 172), with biconvex anterior margin and extending beyond level of st1, lateral margins thickened and posterolateral corners fused with endopodal shields; posterior margin concave; shield not eroded between st1 – st1, bearing three pairs of smooth pointed setae (st1 14 – 16, st2 20 – 22, st3 20 – 22) and two pairs of lyrifissures, lyrifissures iv1 adjacent to setae st1, lyrifissures iv2 between st2 and st3; weak lines near central and posterior margin, anterior area smooth. Seta st4 absent, metasternal pores also on extent of sternal shield but metasternal plates apparently absent. Genito-ventral shield wide, inverted U-shape, strongly tapering posteriorly, 240 – 250 long, 106 – 115 maximum width. Surface of shield has characteristic ornamentation consisting of longitudinal striae; with one pair of simple setae st5 (20 – 22). Anal shield (52 – 60 × 100 – 107 wide) with anterolateral extensions, its anterior without lineate ornamentation, cribrum a narrow band without discernible rows, with a pair of circular lateral gland pores, bearing post-anal seta 8 – 10 long and a pair of para-anal setae 8 – 10 long. Opisthogastric skin with long, narrow and oval metapodal plates (34 – 40 × 8 – 10 wide) and 12 pairs of simple setae 10 – 14 long. Peritreme short (35 – 40), extending from coxa IV to posterior level of coxa II. Peritrematal shields absent, post-stigmatal section conspicuous and narrow, bearing lyrifissure near stigmata and two pore-like structures of post-stigmatal pores.
Gnathosoma — Epistome triangular, irregularly denticulate (Figure 1C). Hypostomal groove with seven rows of denticles, three to six very fine denticles per row (Figure 1D). Hypostome with four pairs of setae, internal posterior hypostomal setae h3 longest. Palp chaetotaxy: trochanter 1, femur 5, genu 6, tibia 11; all palp setae pointed, palp tarsal claw two-tined, dorsodistal edge of palp femur without swelling. Chelicera hyaline, fixed digit of chelicera reduced and edentate, with a low median bulge, pilus dentilis and dorsal lyrifissure absent (Figure 1E); movable digit weakly sclerotised, with one strong terminal hook, cheliceral seta, arthrodial corona and its filaments absent (Figure 1E). Corniculi long, membranous. Lateral malae arms absent (Figure 1D).
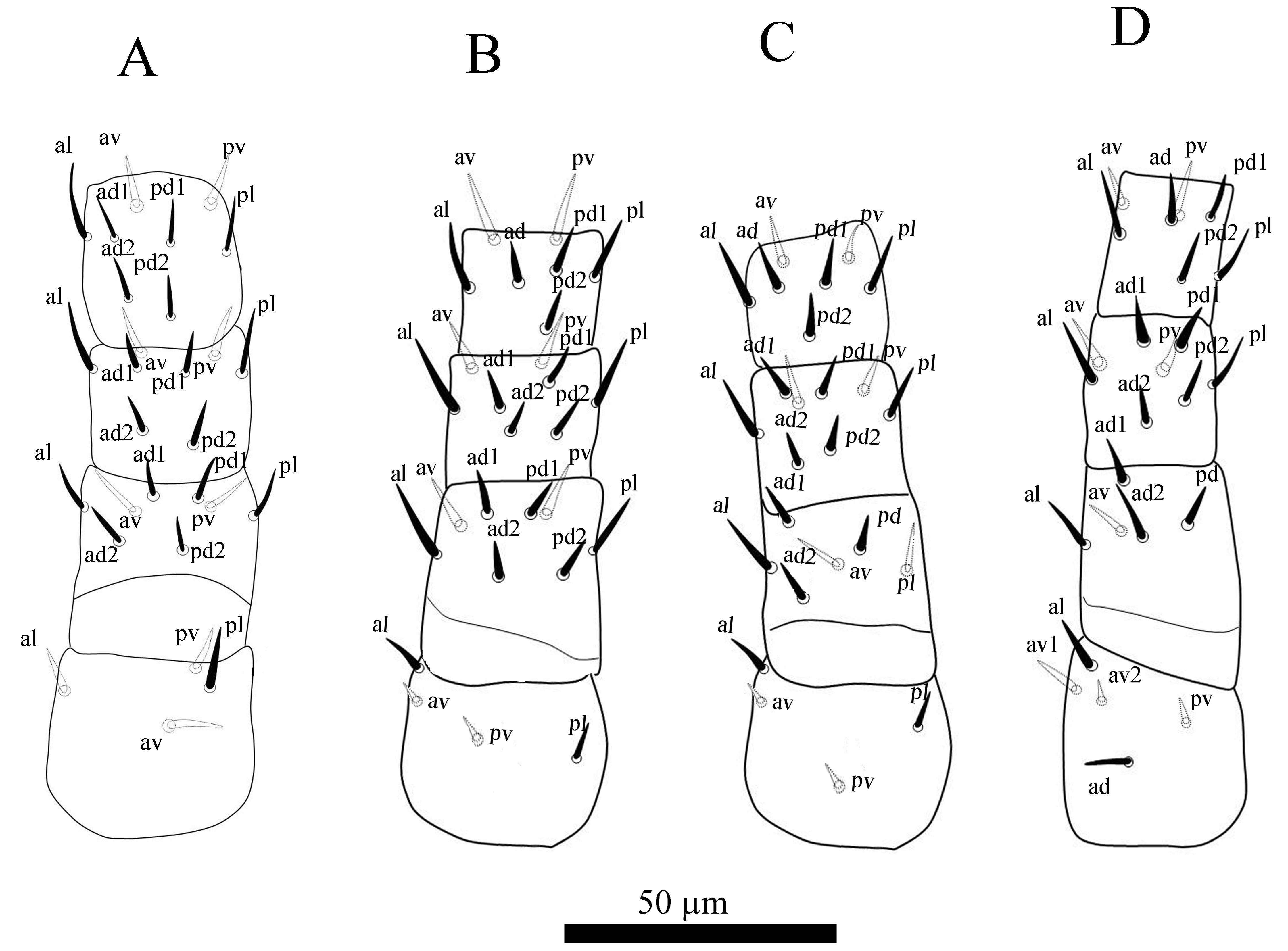
Legs (Figure 2 A–D) — Length of leg I 200 – 210, leg II 190 – 202, leg III 178 – 190, leg IV 190 – 205. Setal counts for legs I–IV: 2–4–8–8–8, 2–4–8–8–7, 2–4–6–8–7, 0–5–5–8–7. Chaetotaxy (both sides examined): Leg I: coxa 0 0/1 0/1 0, trochanter 1 0/1 0/1 1, femur 1 2/1 2/1 1, genu 1 2/1 2/1 1, tibia 1 2/1 2/1 1 (Figure 2A). Leg II: coxa 0 0/1 0/1 0, trochanter 1 0/1 0/1 1, femur 1 2/1 2/1 1, genu 1 2/1 2/1 1, tibia 1 1/1 2/1 1 (Figure 2B). Leg III: coxa 0 0/1 0/1 0, trochanter 1 0/1 0/1 1, femur 1 2/1 1/0 1, genu 1 2/1 2/1 1, tibia 1 1/1 2/1 1 (Figure 2C). Leg IV: coxa 0 0/0 0/0 0, trochanter 1 1/2 0/1 0, femur 1 2/1 1/0 0, genu 1 2/1 2/1 1, tibia 1 1/1 2/1 1 (Figure 2D); all setae smooth. Tarsi II–IV with 18 setae 3 3/2 3/2 3 + mv, md. All pre-tarsi with a pair of thin claws and a membranous ambulacrum.

Genital structures — Insemination ducts opening on posterior margin of coxa III, sacculus indistinct, apparently unsclerotised.
Male. Unknown.
Etymology — The name brachytrichos (Greek brachy, short and trichos, hair) refers to the minute dorsal shield setae.
Remarks — According to the key to species provided by Joharchi et al. (2015), Myrmozercon brachytrichos n. sp. most resembles M. tauricus Trach and Khaustov, 2011. The new species differs from M. tauricus by having: opisthonotal shield with 14 pairs of setae and Z5 in soft skin posterior to shield (versus Z5 in opisthonotal shield in M. tauricus), all setae on shield minute and uniform in length (versus posterior dorsal setae longer than anterior dorsal setae in M. tauricus); epistome triangular, irregularly denticulate (versus smooth in M. tauricus), sternal shield not eroded between st1–st1 (versus eroded between st1–st1 in M. tauricus), setal counts for trochanter I–III (four setae in M. brachytrichos versus five setae in M. tauricus) and for genu IV (eight setae in M. brachytrichos versus seven setae in M. tauricus). Mymrozercon brachytrichos is also very similar to three other species: Myrmozercon eidmanni (Sellnick), Myrmozercon liguricus (Vitzthum) and Myrmozercon minor (Sellnick). The new species differs from M. eidmanni by having no seta on coxa IV and all dorsal shield setae minute and uniform in length (vs coxa IV seta present and seta j1 longer than other body setae in M. eidmanni); it differs from M. liguricus by the slightly reduced fixed cheliceral digit (obsolete fixed digit in M. brachytrichos); and it differs from Myrmozercon minor by having fewer setae on the dorsal shield (34 pairs in M. brachyserose; approximately 31 pairs in M. minor).
Discussion — Myrmozercon brachytrichos n. sp. belongs to the Myrmozercon sensu stricto species group, which has the following character states: leg I femur, genu and tibia 8–8–8; leg II 8–8–7; leg III genu and tibia 8–7; leg IV genu with seven or eight setae, tibia with seven setae; legs short, palp trochanter with one seta; palpcoxal seta sometimes absent; corniculi membranous, dorsal shield smooth, sternal shield an inverted U-shape, setae st4 usually absent and anal shield usually with anterolateral extensions.
Myrmozercon s. str. may have a more intimate association with their hosts because they are almost always collected while they cling onto ants (see Joharchi and Moradi 2013). Almost all have been reported in association with arboreal ants of the genus Crematogaster but nothing is known of their feeding behavior, or any other aspect of their biology. However, their specialized morphology (e.g, edentate chelicerae, short peritremes, corniculi membranous etc.) suggest that Myrmozercon s. str. could be parasitic on its ant hosts, and not simply a commensal in its host’s nests, but this has not been established experimentally.
We gratefully acknowledge Dr. Owen D. Seeman (Queensland Museum, South Brisbane, Queensland, Australia) for reviewing and constructive comments on an earlier version of the manuscript. The authors wish to thank Dr. Farid Faraji (Mitox, Amsterdam, Netherlands) and the other anonymous reviewer for their valuable suggestions.
Babaeian E., Joharchi O., Saboori A. 2013 — A new species of Myrmozercon Berlese (Acari: Mesostigmata: Laelapidae) associated with ant from Iran — Acarologia, 53: 453-460. doi:10.1051/acarologia/20132109 ![]()
Babaeian E., Seraj A., Nemati A. 2014 — Description of a new ant-associated species (Acari: Mesostigmata: Laelapidae) from Iran — Acarologia, 54: 441-448. doi:10.1051/acarologia/20142129 ![]()
Berlese A. 1902 — Specie di Acari nuovi — Zool. Anz., 25: 697-700.
Berlese A. 1903 — Diagnosi di alcuninuove specie di Acari italiani, mirmecofili e liberi — Zool. Anz., 27: 12-28.
Berlese A. 1904 — Illustrazione iconografica degli Acari mirmecofili — Redia, 1: 299–474 + Plates 7-20.
Evans G.O. 1963 — Observations on the chaetotaxy of the legs in the free-living Gamasina (Acari: Mesostigmata) — Bull. Br. Mus. (Nat. Hist.) Zool., 10: 277-303.
Evans G.O., Till W.M. 1979 — Mesostigmatic mites of Britain and Ireland (Chelicerata: Acari-Parasitiformes). An introduction to their external morphology and classification — Trans. Zool. Soc. Lond., 35: 145-270. doi:10.1111/j.1096-3642.1979.tb00059.x ![]()
Ghafarian A., Joharchi O. Jalalizand A., Jalaeian M. 2013 — A new species of Myrmozercon Berlese (Acari: Mesostigmata: Laelapidae) associated with ants from Iran — ZooKeys, 272: 21-28.
Hunter P.E., Hunter C.A. 1963 — The genus Myrmonyssus with descriptions of two new species (Acarina: Laelaptidae) — Acarologia, 5: 335-341.
Joharchi O., Babaian E., Seeman O.D. 2015 — Review of the genus Myrmozercon Berlese (Acari: Laelapidae), with description of a new species from Iran — Zootaxa, 3955(4): 549-560.
Joharchi O., Halliday B., Saboori A., Kamali K. 2011 — New species and new records of mites of the family Laelapidae (Acari: Mesostigmata) associated with ants in Iran — Zootaxa, 2972: 22-36.
Joharchi O., Moradi M. 2013 — Review of the genus Myrmozercon Berlese (Acari: Laelapidae), with description of two new species from Iran — Zootaxa, 3686: 244-254.
Karawajew W. 1909 — Myrmekophilen aus Transkaspien — Russkoe Entomologicheskoe Obozrenie, 9: 227-237.
Lindquist E.E., Evans G.O. 1965 — Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina (Acarina: Mesostigmata) — Mem. Ent. Soc. Can., 47: 1-64. doi:10.4039/entm9747fv ![]()
Rosario R.M.T., Hunter P.E. 1988 — The genus Myrmozercon Berlese, with descriptions of two new species (Acari: Mesostigmata: Laelapidae) — J. Parasitol., 74: 466-470. doi:10.2307/3282057 ![]()
Sellnick M. 1941 — Milben von Fernando Poo — Zool. Anz., 136: 221-228.
Shaw M.D., Seeman O.D. 2009 — Two new species of Myrmozercon (Acari: Laelapidae) from Australian ants (Hymenoptera: Formicidae) — Zootaxa, 2025: 43-55.
Trach V.A., Khaustov A.A. 2011 — A myrmecophilous mite Myrmozercon tauricus sp.n. of the family Laelapidae (Acari, Mesostigmata) from Ukraine — Vestn. Zool., 45: 23-27.
Ueckermann E.A., Loots G.C. 1995 — A new laelapid genus and species (Acari: Parasitiformes: Laelapidae) from Yemen — Afr. Entomol., 3: 35-38.
Vitzthum H.G. 1930 — Ein Ameisengast (Acar.) — Mitt. Deut. Entomol. Gesell. Ber., 6: 89-94.
Walter D.E. 2003 — A new mite from an arboreal ant (Formicidae: Polyrachis sp.): Myrmozercon iainkayi n. sp. (Mesostigmata: Laelapidae) — Int. J. Acarol., 29: 81-85. doi:10.1080/01647950308684325 ![]()
Walter D.E., Krantz G.W. 2009 — Collecting, rearing and preparing specimens. In: Krantz G.W., Walter D.E. (eds) A Manual of Acarology, Third Edition, Texas Tech University Press, Lubbuck, Texas, pp. 83-95.



2016-11-25
Date accepted:
2017-02-03
Date published:
2017-07-06
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2017 Joharchi, Omid; Arjomandi, Elham and Trach, Viacheslav A.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)