



Demographic analysis of sublethal effects of spiromesifen on Neoseiulus californicus (Acari: Phytoseiidae)
Ghadim Mollaloo, Maryam1 ; Kheradmand, Katayoon2 ; Sadeghi, Reza3 and Talebi, Ali Aasghar4
1Department of Entomology and Plant Pathology, College of Aboureyhan, University of Tehran, Pakdasht, Iran
2✉ Department of Entomology and Plant Pathology, College of Aboureyhan, University of Tehran, Pakdasht, Iran
3Department of Entomology and Plant Pathology, College of Aboureyhan, University of Tehran, Pakdasht, Iran
4Department of Entomology, Faculty of Agriculture, Tarbiat Modares University, Tehran, Iran
2017 - Volume: 57 Issue: 3 pages: 571-580
https://doi.org/10.24349/acarologia/20174173Keywords
Abstract
Phytoseiidae is a family of mites which feed on thrips and other mite species. Because of their usefulness as biological control agents, interest in Phytoseiids has steadily increased over the past century (Waite 1998; Van Lenteren 2000). Phytoseiids can postpone or delay egg production during periods where preys are scarce. This allows them to have a longer lifespan and likely serves as an adaptation to environments where prey availability is variable (McMurtry et al. 1970). The predatory mite Neoseiulus californicus (McGregor) is used commercially around the world to control the two-spotted spider mite and several other economically important mites on avocado, citrus, dwarf hops, grapes, raspberries, roses and other ornamentals, strawberries, and several vegetable crops (Castagnoli and Simoni 1991; Gotoh et al. 2004).
This predatory mite develops faster when consuming the two-spotted spider mite, Tetranychus urticae Koch, than when consuming other prey sources (Jolly 2000). If two-spotted spider mite population is already high, it may be necessary to spray a miticide before releasing N. californicus (Calvo et al. 2012). Spiromesifen is a novel non-systemic insecticide/acaricide belonging to the new chemical class of spirocyclic phenyl-substituted tetronic acids. It is especially active against whiteflies (Bemisia spp.and Trialeurodes spp.) after foliar application and is highly active against Tetranychus spp. as an acaricide in many cropping systems such as cotton, vegetables and ornamentals (Nauen et al. 2002). But despite of its widespread usage, Kaplan et al., 2012, reported that spirodiclofen can be harmful for N. californicus at field application dosage (Kaplan et al. 2012). epMost toxicological studies have focused on measuring the lethal effects of this pesticides on predatory mites (Raudonis 2006; Bostanian et al. 2009; Duchovskienė et al. 2009; Kaplan et al. 2012). But it may be a partial measure of their deleterious effects and so sublethal effects can be very subtle and affect populations at concentrations lower than the traditional concentration and can complete our information about its sublethal effects on physiology and behavior of arthropods (Stark and Banks 2003; Desneux et al., 2007). For this purpose many researchers have used Probit regression line for depicting relationship between probit of mortality and log concentration of pesticide following 24 h of exposure. In addition, evaluation of pesticide effects based solely on females would not exhibit the overall impacts of pesticides, so it's necessary to conduct the specifying sublethal effects on biology of both sexes of the mite (Alinejad et al. 2014). Therefore, in this study we apply the age-stage, two-sex life table approach to analyze the life history data and life parameters of N. californicus to incorporate the variable developmental rates among individuals and the male population.epSo, the aim of this work was to evaluate the sublethal effect of spiromesifen on the survival and reproduction of N. californicus to determine the compatibility of the mentioned acaricide as an effective pesticide with N. californicus can be used to manage T. urticae population.
The colony of N. californicus was provided by Giah Bazr Alvand Company, agent of Koppert Company in Tehran and reared in the laboratory on kidney bean plants (Phaseolus vulgaris L) infested with T. urticae. The colony of T. urticae was collected from infested greenhouses located in Varamin (south eastern of Tehran). The rearing arenas were constructed according to Abou-Setta and Childers (1987) method with some changes. A 1 cm diameter hole was drilled in the center of a 9 cm diameter plastic Petri dish. The first Petri dish was sealed using glue over another 9 cm diameter Petri dish, which was used to hold water. Afterward, a 2 mm diameter hole was created in the upper part of the side wall of the second Petri dish for filling it with water using a squeeze bottle. A narrow wick made from filter paper was inserted to the second Petri dish through the hole of the first Petri dish. The wick supplies water for keeping a bean leaf fresh in the first Petri dish. Then a thin cotton wool layer was located at the bottom of the first Petri dish and was covered with filter paper. An upturned bean leaf was placed on the filter paper and surrounded with saturated cotton to prevent escape of the mites. The rearing arena was covered with a ventilated lid.
A commercial formulation of spiromesifen (IUPAC name: [2-oxo-3-(2,4,6-trimethylphenyl)-1-oxaspiro[4-4]non-3-en-4-yl]3,3-dimethylbutanoate) (Oberon®, Bayer CropScience) was used in the present study. The response of N. californicus adults to various concentrations of spiromesifen was assessed using the leaf dip bioassay method (Helle and Overmeer 1985). The experimental arena was the same as the rearing arena but using 6 cm diameter Petri dish. Freshly cut bean leaf discs of 4 cm diameter were dipped into various spiromesifen solutions for 15 seconds and air dried for about 3 hours. The control leaf discs were dipped into distilled water. Each leaf disc was transferred into the experimental arena. Afterward, forty same age (less than 24-h-old) adult mites (male and female, 20:20) of the same age (less than 24-h-old) were placed on the treated leaf discs for each concentration. The main doses obtained from preliminary tests was performed in four replicates to get sublethal concentrations. Mortality of the mites was assessed after 24 hour of exposure. Abbott's formula was used to estimate the natural mortality (Abbott 1925).
After treatment of bean leaf discs with sublethal concentrations of spiromesifen (LC5, LC10 and LC15), same aged (24 h old) females were placed on the discs. After 24 hours, the surviving females were separately introduced onto untreated bean leaf discs (20 mm in diameter). Subsequently, after 24 hours, the eggs laid by the treated and untreated females in each experimental arena were saved. All saved eggs were checked daily and development time and survival were recorded. Each predatory mite was fed with four to six immature stages of spider mite daily and enough corn pollen every week. Newly emerged females were coupled with a male for mating. The daily survival of each adult and fecundity of females were recorded and population parameters were calculated.
The sublethal concentrations (LC5, LC10 and LC15) were determined using a probit procedure. The raw data of the life table parameters were analyzed according to the theory of age-stage, two-sex life table (Chi and Liu 1985) by using a user-friendly computer program, TWOSEX-MS Chart (Chi 2012). The age-stage specific survival rate (Sxj), age-specific survival rate (lx), age-specific fecundity (mx), and population parameters were calculated accordingly.
Intrinsic rate of increase was estimated using the iterative bisection method from the Euler-Lotka formula:
Σx=0 ∞ e-r(x+1) lxmx = 1,
with age indexed from 0 (Goodman, 1982). The mean generation time is defined as the length of time that a population needs to increase to R0-fold of its size (i.e. e rT =R 0 or λT = R 0) at the stable age-stage distribution and is calculated as T = (lnR 0)/r. Paired bootstrap method in Two-Sex, Age-Stage life table software (2015 version) was used to estimate the means and standard errors with 10,000 replications (Huang and Chi 2013). Bioassay data obtained were subjected to one-way ANOVA (P≤0.05) after checking for normality. Means were compared by Tukey’s test, admitting significant differences at P≤0.05. SPSS 19.0 software was used for mean comparisons.
The LC50 value for treatments with spiromesifen, and its 95% confidence interval, calculated from probit regression, was estimated 53.779 ppm. No mortality was recorded in the controls (Table 1). The sublethal concentrations for N. californicus were 5. 834 ppm, 9.529 ppm and 13.267 ppm for LC5, LC10 and LC15, respectively.


The effects of different concentrations of spiromesifen on developmental time, longevity and total life span of offspring from the treated females are presented in Table 2. The time required for N. californicus eggs to hatch was 1.1 and 1.21 days for the untreated males and females, respectively. Sublethal concentration was not affected egg incubation time either male or female. This nontoxic effect of spiromesifen showed in larval, protonymph and deutonymph periods on both sex. The longevity and total life span of treated male (df =3; F=302.42; P=0.0004) and female (df=3; F=639.13; P=0) treated with LC15 concentration of spiromesifen was significantly different from control and other sublethal concentration.


No significant differences between all experimental doses and control were scored in adult pre-oviposition and total pre-oviposition period. Treatment with LC5 sublethal concentration of spiromesifen showed no significant change in total fecundity about per female of N. californicus nearly like control treatment. By increasing in concentration at LC10, harmful effect of spiromesifen showed relatively in oviposition duration but non-effect in pre- oviposition period. In other word LC10 demonstreted an intermediate concentration between non-effecting dosage (LC5) and effecting dosage (LC15). Applying LC15 and LC10 concentrations caused significant reduction in mites total fecundity (Table 3).

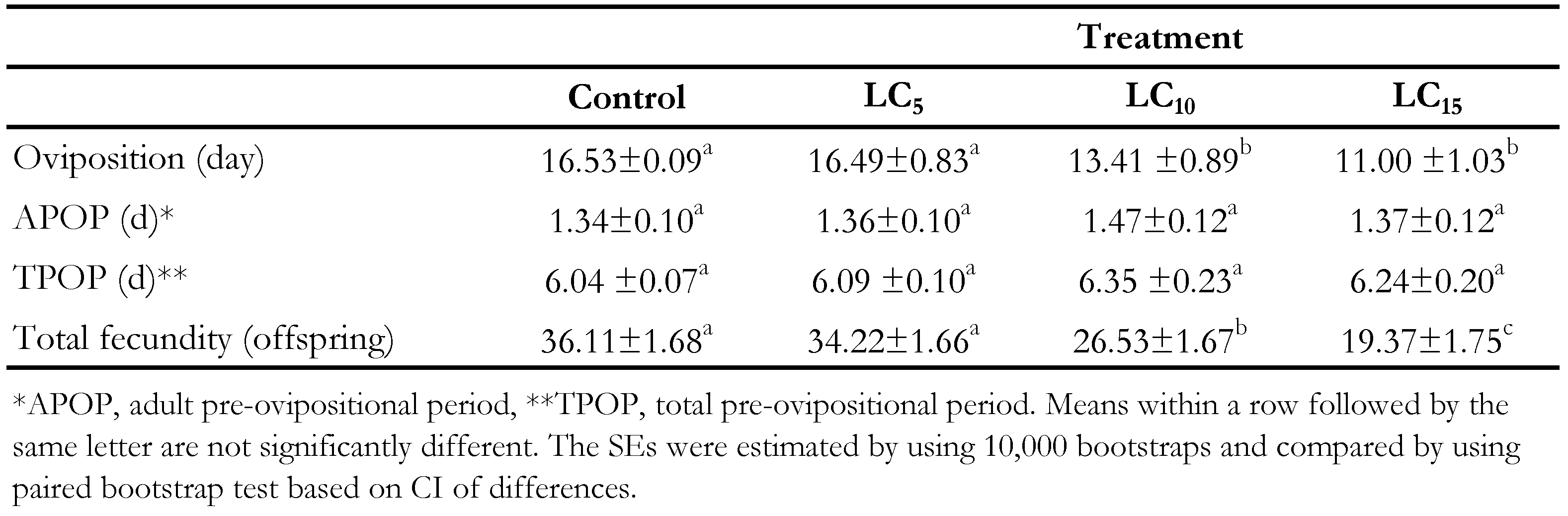
The life table parameters of offspring related to the treated females are demonstrated in Table 4. The intrinsic rate of increase (r) was varied between 0.17 and 0.24 day-1 that the highest one was related to the control treatment. So, the (r) gradually decreased by increasing the concentrations of spiromesifen. The highest and lowest finite rate of increase (λ) was observed in control and LC15 concentrations which showed significant difference. The net reproductive rate ( R 0) significantly decreased by increasing the concentrations of spiromesifen specially in LC10 and LC15 treatments. The gross reproductive rates ( GRR) was 26.71 and 14.72 offspring in control and LC15 respectively that showed its reduction by increasing concentration of acaricide. Although the mean generation time ( T) in control and other treatments, had not significant differences.

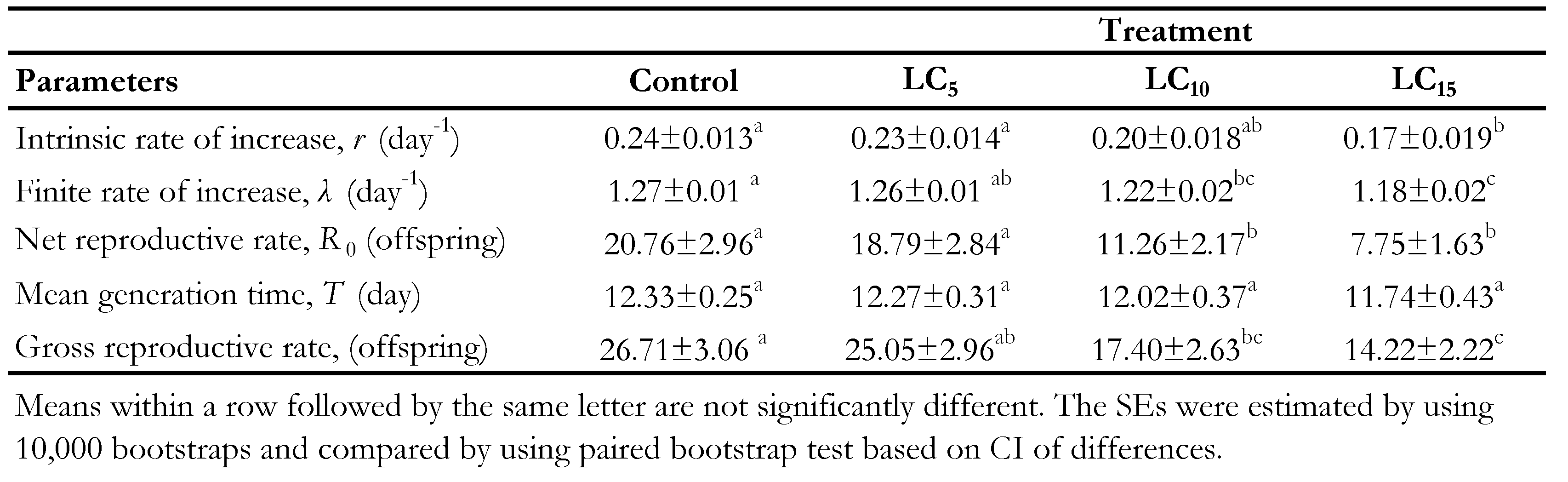
Figure 1 Shows the daily and age-stage survival curves of both untreated and treated individuals of N. californicus with sublethal concentrations of spiromesifen . The total lifetime for the untreated mites was 38 days, while it was 38, 31 and 28 days for LC 5, LC 10 and LC15 treatments, respectively. In addition, examination of lx in treated mites revealed more mortality in immature stages in contrast to control. The age-stage survival rate (Sxj) illustrates the probability that a new born will survive to age x and stage j (Figure 2). There are significant overlaps between curves of different stages. and different sublethal concentrations. The highest female and male survival rate was observed at control treatment and effect of LC5 showed same trend like survival rates of control. By increasing in concentration (LC10 and LC15), survival rates declined in compare with what was in control.The maximum values of mx were approximately 1.68 eggs/female/day for the mites treated by LC5 sublethal concentration, which was occurred on day 9 of the life span (Figure 3). In comparison, maximum value of 1.75 eggs/female/day was observed on day 12 of the life span for the untreated mites. But maximum values of mx were approximately 1.4 and 1.2 eggs/female/day which was occurred on days 10 and 18 of the life span for the mites contacted with LC10 and LC15 concentrations, respectively.

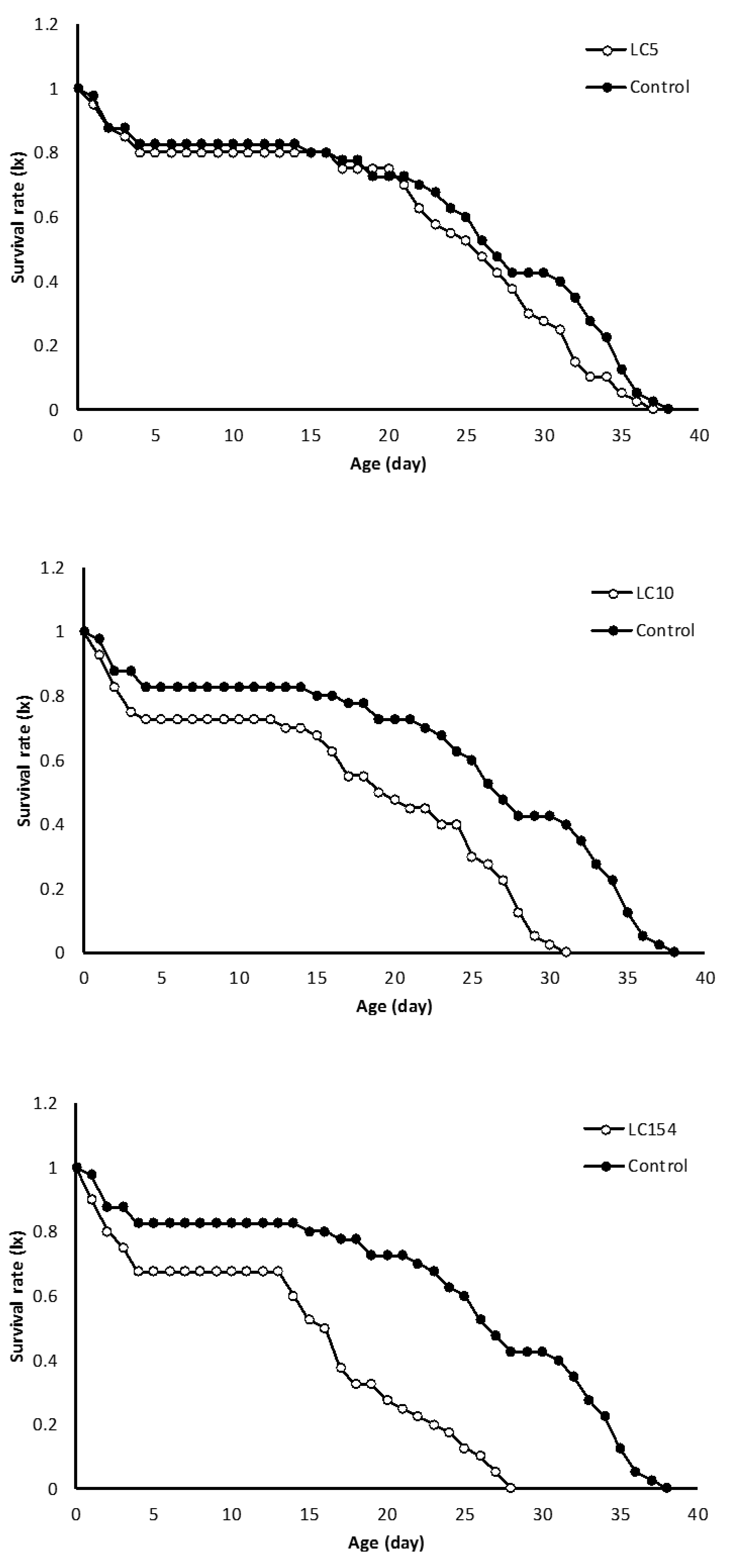

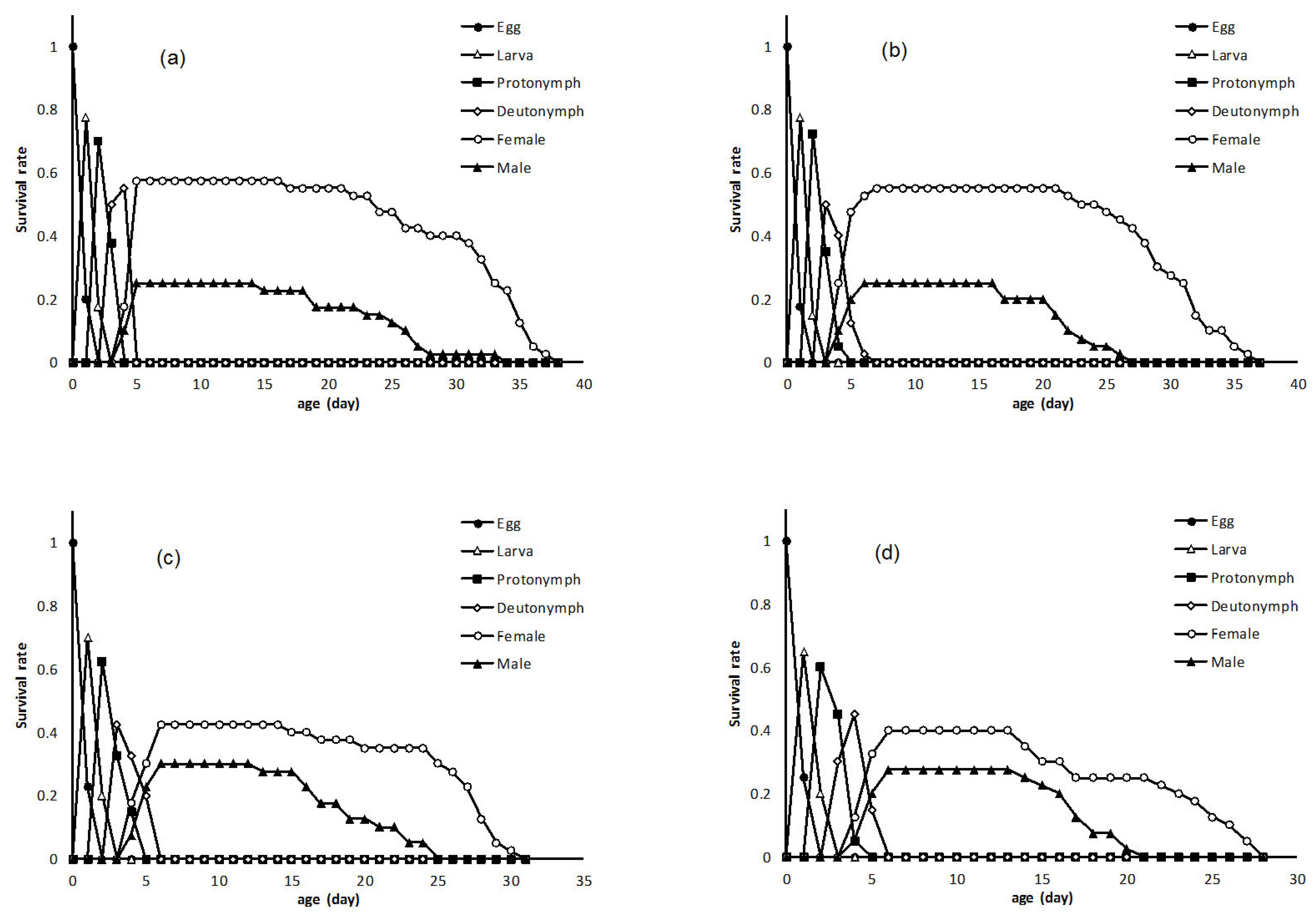


It has been reported that spiromesifen acts on mite and whitefly development probably by interfering with lipid biosynthesis (Nauen et al. 2002). There is no information on the sublethal effects of spiromesifen on biological characteristic of N.californicus, including development time, fecundity, mortality at different life stages and adult emergence. In present study, although the concentrations (LC5, LC10, LC15) used were lower than what was recommended rate for two spider mite. Moreover, application of sublethal concentrations of spiromesifen had no significant adverse effects on all immature stages of both sexes of N. californicus. Our results are consistent with Hamedi et al. (2010) that mentioned application of sublethal concentrations of abamectin had no significant effect on larval period of Phytoseiulus plumifer.
In spite of no significant effect of sublethal concentrations of spiromesifen on premature stages, this acaricide decreased adult longevity and total life span of both sexes along with increasing of dosages. This result was confirmed in bioassay related to Alinejad et al. (2014) concerning effects of sublethal concentrations of fenazaquin on A. swirskii Although application of sublethal concentrations of spirodiclofen had no negative effect on these parameters of A. swirskii (Alinejad et al. 2016). Moreover, Hamedi et al. (2011) had shown the same results like ours on P. plumifer by increasing of abamectin concentrations as well as they had confirmed this subject about fenpyroximate, previously (Hamedi et al. 2009).
LC10 and LC15 sublethal concentrations caused reduction significantly in the total fecundity period of N. californicus. Also Park et al. (2011) and Alinejad et al. (2014) noted the similar trend for pyridaben and fenazaquin, respectively. In this manner Ibrahim and Yee (2000) reported that the exposure to sublethal concentration of abamectin reduced the mean fecundity per female of Neoseiulus longispinosus (Evans) to almost half of the untreated females. But they had inconsistent conclusion about application of sublethal concentrations of spirodiclofen. Differences in susceptibility of phytoseiid species and tested acaricides could be responsible for the conflicting results.epIn addition to research on the effects of pesticides on predators, Marcic et al. (2010) showed that Spiromesifen at concentrations of 180, 18 and 1.8 mg/l significantly reduced fecundity, fertility and population growth of T. urticae . In contrary, Barati and Hejazi (2015) indicated that all neonicotinoids tested increased the fecundity of T. urticae because of changes in physiology of the mites. So the use of acaricides against mites in the presence of predators requires necessary precautions. Consequently, use of spiromesifen in crops should be limited while this predator is important for biological control to get a better result. It is necessary to pay attention to the direct toxicity of spiromesifen on N.californicus for considering this accaricide in IPM program. The sublethal effects of pesticides on life table parameters of phytophagous and predatory mites were previously reported by several researchers (Alinejad et al. (2014), Ghaderi et al. (2013), Hamedi et al. (2010), Ibrahim and Yee (2000) and Marcic (2007) demonstrated decline trend for population parameters of A.swirski, P. persimilis, P. plumifer , N.longipinosus and T.urticae , when treated by sublethal concentrations of fenazaquin, fenpyroximate, abamectin and spirodiclofen, respectively).Our results revealed that LC15 sublethal concentration of spiromesifen had significant effect on population parameters as confirmed in previous parts and are corroborated by Ibrahim and Yee (2000), Alinejad et al. (2014), Hamedi et al. (2011) and Ghaderi et al. (2013).
The age specific fecundity and survival curves demonstrated that sublethal concentrations of spiromesifen caused significant reduction in these parameters of N. californicus, to compare with the control. Like the study of Alinejad et al. (2014) and Ibrahim and Yee (2000) in which the sublethal concentrations of fenazaquin and abamectin were examined on A. swirskii and N. longispinosus respectively.
Conclusively, this study has revealed that treating N. californicus with increasing lethal concentration of spiromesifen decreased fecundity and population numbers of N. californicus under laboratory conditions. However, it's necessary to consider the direct toxicity of spiromesifen on N. californicus in IPM programs. Final judgment on the effect of high sublethal concentration of spiromesifen on N. californicus fecundity and life table parameters should be postponed until the results are obtained from semi-field and field trials. If results are similar, the use of these chemicals in IPM programs requires necessary precautions.
We appreciate University of Tehran for the financial support of this study.
Abbott W.S. 1925 — A method of computing the effectiveness of an insecticide — J. Econom. Entomol., 18: 265–267. doi:10.1093/jee/18.2.265a ![]()
Abou Setta M.M., Childers C.C. 1987 — A modified leaf arena technique for rearing phytoseiid or tetranychid mites for biological studies —Fla. Entomol., 70: 245–248. doi:10.2307/3495156 ![]()
Alinejad M., Kheradmand K., Fathipour Y. 2014 — Sublethal effects of fenazaquin on life table parameters of the predatory mite Amblyseius swirskii (Acari: Phytoseiidae) — Exp. Appl. Acarol., 64: 361–373. doi:10.1007/s10493-014-9830-y ![]()
Alinejad M., Kheradmand K., Fathipour Y. 2016 — Assessment of sublethal effects of spirodiclofen on biological performance of the predatory mite, Amblyseius swirskii — Syst. Appl. Acarol., 21: 375-384.
Barati R., Hejazi M.J. 2015 — Reproductive parameters of Tetranychus urticae (Acari: Tetranychidae) affected by neonicotinoid insecticides — Exp. Appl. Acarol., 66: 481-489. doi:10.1007/s10493-015-9910-7 ![]()
Bostanian N. J., Beudjekian S., McGregor E., Racette G. 2009 — A modified excised leaf disc method to estimate the toxicity of slow and fast acting reduced risk acaricides to mites — J. Economi. Entomol., 102: 2084–2089. doi:10.1603/029.102.0610 ![]()
Calvo F.J., Bolckmans K., Belda J.E. 2012 — Biological control-based IPM in sweet pepper greenhouses using Amblyseius swirskii (Acari: Phytoseiidae) — Biocontrol Sci. Techn., 22: 1398-1416. doi:10.1080/09583157.2012.731494 ![]()
Castagnoli M., Simoni S. 1991 — Influenza della temperature sullincremento delle popolazioni di Amblyseius californicus (McGregor) (Acari: Phytoseiidae) — Redia, 74: 621-640.
Chi H. 1988 — Life table analysis incorporating both sexes and variable development rates among individuals — Environ. Entomol., 17: 26-34. doi:10.1093/ee/17.1.26 ![]()
Chi H., Liu H. 1985 — Two new methods for the study of insect population ecology — Bulletin of the Institute of Zoology Academia Sinica, 24: 225–240.
Chi H. — TWOSEX-MSChart: a computer program for the age-stage, two-sex life table analysis — [20 June 2012]. Available from: http://140.120.197.173/Ecology/. National Chung Hsing University, Taichung Taiwan.
Desneux, N., Decourtye, A., Delpuech, J.M. 2007 — The sublethal effects of pesticides on beneficial arthropods — Annual Review of Entomology, 52: 81–106. doi:10.1146/annurev.ento.52.110405.091440 ![]()
Duchovskienė L., Raudonis L., Karklelienė R., Starkutė R. 2009 — Toxicity of insecticides to predatory mite Phytoseuilus persimilis in cucumber. Scientific Works of the Lithuanian Institute of Horticulture and Lithuanian University of Agriculture — Sodininkyst Ir Daržininkyst., 28: 41–46.
Ghaderi S., Minaei K., Kavousi A., Akrami M.A., Aleosfoor M., Ghadamyari M. 2013 —Demographic analysis of the effect of Fenpyroximate on Phytoseiulus persimilis Athias-Henriot (Acari: Phytoseiidae) — Entomol. Gen., 34: 225–233. doi:10.1127/entom.gen/34/2013/225 ![]()
Gotoh T., Yamaguchi K., Mori K. 2004 — Effect of temperature on life history of the predatory mite Amblyseius californicus (Acari: Phytoseiidae) — Exp. Appl. Acarol., 32: 15-30. doi:10.1023/B:APPA.0000018192.91930.49 ![]()
Hamedi N., Fathipour Y., Saber M. 2009 — Sublethal effects of fenpyroximate on life table parameters of the predatory mite extitPhytoseius plumifer
— Biocontrol, 55: 271–278.Hamedi N., Fathipour Y., Saber M. 2010 — Sublethal effects of abamectin on the biological performance of the predatory mite, Phytoseius plumifer (Acari: Phytoseiidae) — Exp. Appl. Acarol., 53: 29–40. doi:10.1007/s10493-010-9382-8 ![]()
Hamedi N., Fathipour Y., Saber M., 2011 — Sublethal effects of abamectin on the biological performance of the predatory mite, Phytoseius plumifer (Acari: Phytoseiidae) — Exp. Appl. Acarol., 53:.29–40.
Helle W., Overmeer W.P.J. 1985 — Toxicological test methods — In: Helle W., Sabelis M.W. (Eds). Spider Mites. Their Biology, Natural Enemies and Control: Elsevier. pp. 391–395.
Huang Y. B., Chi H. 2013 — Life tables of Bactrocera cucurbitae (Diptera: Tephritidae): with an invalidation of the jackknife technique — J. Appl. Entomol., 137: 327–339. doi:10.1111/jen.12002 ![]()
Ibrahim Y.B., Yee T.S. 2000 — Influence of sublethal exposure to abamectin on the biological performance of Neoseiulus longispinosus (Acari: Phytoseiidae) — J. Econom. Entomol., 93: 1085–1089. doi:10.1603/0022-0493-93.4.1085 ![]()
Jolly R.L. 2000 — The predatory mite Neoseiulus californicus: its potential as biological control agent for the fruit tree red spider mite, Panonychus ulmi — The BPC Conference at Brighton, Pest and Control, 1: 487–490.
Kaplan P., Yorulmaz S., Ay R. 2012 — Toxicity of insecticides and acaricides to the predatory mite Neoseiulus californicus (McGregor) (Acari: Phytoseiidae) — Int. J. Acarol., 38: 699–705. doi:10.1080/01647954.2012.719031 ![]()
Marcic D. 2007 — Sublethal effects of spirodiclofen on life history and life-table parameters of two-spotted spider mite (Tetranychus urticae) — Exp. Appl. Acarol., 42: 121–129. doi:10.1007/s10493-007-9082-1 ![]()
Marcic D., Ogurlic I., Mutavdzic S., Peric P. 2010 — The effects of spiromesifen on life history traits and population growth of two-spotted spider mite (Acari: Tetranychidae) — Exp. Appl. Acarol., 50: 255–267. doi:10.1007/s10493-009-9316-5 ![]()
McMurtry J.A., Huffaker, C.B., Van de Vrie M. 1970 — Ecology of tetranychid mites and their natural enemies: a review. I. Tetranychid Enemies: their biological characters and the impact of spray practices — Hilgardia, 40: 331–390.
Nauen R., Bretschneider T., Bruck E., Elbert A., Reckmann U., Wachendorff U., Tiemann R. 2002 — A novel compound for whitefly and spider mite control — BCPC Conference, Pests and Diseases., 1: 39–44.
Park J.J., Kim M., Lee J.H., Shin K.I., Lee S.E., Kim J.G., Cho K. 2011 — Sublethal effects of fenpyroximate and pyridaben on two predatory mite species, Neoseiulus womersleyi and Phytoseiulus persimilis (Acari, Phytoseiidae) — Exp. Appl. Acarol., 54: 243–259. doi:10.1007/s10493-011-9435-7 ![]()
Raudonis L. 2006 — Comparative toxicity of spirodiclofen and lambdacihalotrin to Tetranychus urticae, Tarsonemus pallidus and predatory mite Amblyseius andersoni in a strawberry site under field conditions — Agron. Research, 4: 317–322.
Stark J.D., Banks J.E. 2003 — Population-level effects of pesticides and other toxicants on arthropods — Ann. Rev. Entomol., 48: 505–519. doi:10.1146/annurev.ento.48.091801.112621 ![]()
Van Lenteren J.C. 2000 — A Greenhouse without Pesticides: Fact of Fantasy — Crop Protection, 19: 375-384. doi:10.1016/S0261-2194(00)00038-7 ![]()
Waite G.K. 1998 — Integrated control of Tetranychus urticae in strawberries in Southest Queensland — Exp. Appl. Acarol., 5: 23-32. doi:10.1007/BF02053814 ![]()



2016-07-03
Date accepted:
2016-12-21
Date published:
2017-05-15
Edited by:
Bonafos, Romain

This work is licensed under a Creative Commons Attribution 4.0 International License
2017 Ghadim Mollaloo, Maryam; Kheradmand, Katayoon; Sadeghi, Reza and Talebi, Ali Aasghar
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)