



The tropics as ancient cradle of oribatid mite diversity
Pachl, Patricia1 ; Lindl, Ann Christin2 ; Krause, Alena3 ; Scheu, Stefan4 ; Schaefer, Ina5 and Maraun, Mark6
1Georg August University Göttingen, JFB Institute of Zoology and Anthropology, Berliner Str. 28, 37073 Göttingen, Germany
2Georg August University Göttingen, JFB Institute of Zoology and Anthropology, Berliner Str. 28, 37073 Göttingen, Germany
3Georg August University Göttingen, JFB Institute of Zoology and Anthropology, Berliner Str. 28, 37073 Göttingen, Germany
4Georg August University Göttingen, JFB Institute of Zoology and Anthropology, Berliner Str. 28, 37073 Göttingen, Germany
5Georg August University Göttingen, JFB Institute of Zoology and Anthropology, Berliner Str. 28, 37073 Göttingen, Germany
6✉ Georg August University Göttingen, JFB Institute of Zoology and Anthropology, Berliner Str. 28, 37073 Göttingen, Germany
2017 - Volume: 57 Issue: 2 pages: 309-322
https://doi.org/10.1051/acarologia/20164148Keywords
Abstract
Species richness of many animal and plant taxa increases from temperate towards tropical regions; a phenomenon known as the latitudinal biodiversity gradient (LBG) (Rohde 1992, Roy et al. 1998, Willig et al. 2003, Hillebrand 2004, Arita & Vázquez-Domínguez 2008, Jablonski et al. 2013, Moreau & Bell 2013, Mannion et al. 2014, Rolland et al. 2015). Explanations for higher species diversity in the tropics as compared to high latitude biomes include biotic and abiotic factors, as niches are smaller in mature communities and climate fluctuations are less pronounced. Additionally, area size may contribute to the higher species richness in the tropics as the area of tropical regions exceeds e.g., that of temperate regions (Chown & Gaston 2000). Moreover, the tropics were not glaciated during the Pleistocene, allowing more continuous radiation in low latitude biomes. Three models focus on the importance of evolutionary time for the high species diversity in tropical regions. The "cradle model" proposes higher speciation rate for tropical regions (Stebbins 1974, Stenseth 1984, Mittelbach et al. 2007). In contrast, the "museum model" assumes lower extinction rates in tropical than in other regions (Stebbins 1974). The "out of the tropics model" combines these two models by postulating higher speciation rates and lower extinction rates in the tropics, with species mainly originating in the tropics and spreading to high latitude regions (Jablonski 1993; Jablonski et al. 2006). The LBG is well investigated in birds, reptiles, frogs, freshwater fish, termites, ants, beetles and trees (Rohde 1992, Hillebrand 2004, Abensperg-Traun & Steven 1997; Wiens et al. 2006). Species with contrasting diversity patterns (i.e., taxa with more species in high latitude regions) such as penguins, seals and sea lions (Proches 2001) typically only occur in cold climates. However, the LBG has been little studied in below-ground living animal taxa. Oribatid mites (Acari, Oribatida) are species-rich, highly abundant microarthropods that occur in soils and decomposing organic matter all over the world. About 10,000 species are described (Subías 2004) but more than 100,000 may exist (Walter & Proctor 2013). Oribatid mites are divided into six major phylogenetic groups (Palaeosomata, Enarthronota, Parhyposomata, Mixonomata, Desmonomata, Brachypylina) (Subías 2004, Norton & Behan-Pelletier 2009), each including cosmopolitan, tropical and temperate species. Patterns of oribatid mite species diversity only in part match the LBG as species diversity increases from boreal to temperate regions but then little further towards tropical regions (Maraun et al. 2007). Generally, oribatid mites are an ancient taxon with the first generally accepted fossil documented from the Devonian (Norton et al. 1988, Subías & Arillo 2002), but their origin may date back to Cambrian or even Precambrian times (Schaefer et al. 2010) when the Earth climate was at least episodically cold as indicated by the magnitude of sea level changes (Miller et al. 2005, Runkel et al. 2010). Due to the ancient age of the group the region where they radiated cannot be delineated easily and ascribed e.g., to tropical or temperate regions, as they were relocated due to continental drift (Hammer & Wallwork 1979). The long-term evolution of oribatid mites potentially facilitated parallel radiations in different climatic regions and continents and thereby convergent evolution of traits (Pachl et al. 2012).
Investigating global distribution patterns in a phylogenetic context is likely to improve understanding of factors that shaped the evolution and radiation of oribatid mites. We constructed a molecular phylogeny of oribatid mites based on two ribosomal markers (partial 18S and 28S rDNA). We used species from each of the main taxonomic groups of oribatid mites and included 28 tropical, 26 temperate and 22 cosmopolitan species. Subsequently, we mapped the current biogeographical distribution of the respective species on the phylogenetic tree and reconstructed the ancestral biogeographic distribution to evaluate which region (temperate, tropical) is the ancestral one. According to the 'museum hypothesis', tropical species should be phylogenetically early-derived with basal position in the phylogenetic tree. In contrast, a scattered and derived distribution of tropical species would support the 'cradle hypothesis'. We hypothesized that tropical regions are regions where oribatid mite speciation took place early in evolution, which would support the "tropics as museum" hypothesis.
In total, 76 oribatid mite taxa from the six major groups and two outgroup taxa (Parasitiformes) were included in this study. In total 69 oribatid mites were identified at species level and seven at genus level. Species were collected from tropical montane rain forests in southern Ecuador (Illig et al. 2010), from temperate forests in central Germany (Erdmann et al. 2012), from the United States of America (obtained from Roy A. Norton, Syracuse, USA) and from a number of sites all over the world (Table 1). Some sequences were available from NCBI, but most were generated for this study. Animals were extracted by heat (Kempson et al. 1963) and determined using Balogh & Balogh (1988, 1990, 2002) and Weigmann (2006). The 18S and 28S rDNA sequences of outgroup taxa (Amblyomma sphenodonti, Ixodidae, and Opilioacarus (Neocarus) texanus, Opilioacaridae) were taken from the NCBI databank.
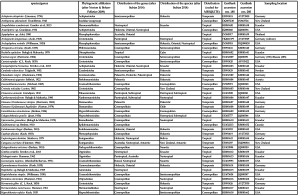

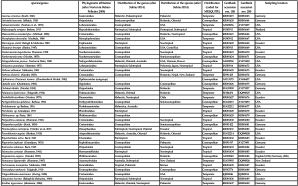
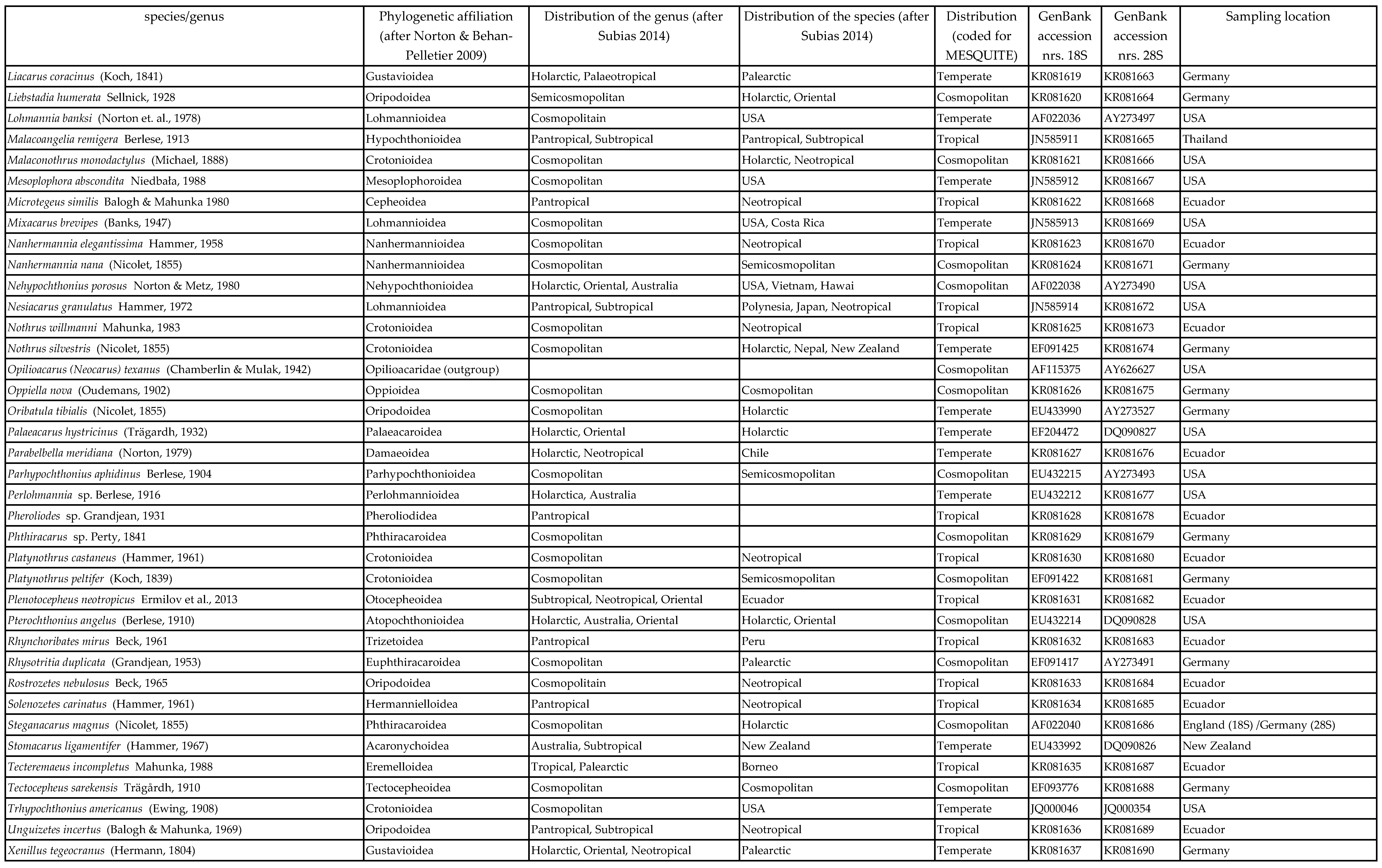
Species were assigned to taxonomic groups according to the classification of Norton & Behan-Pelletier (2009). Distribution patterns (i.e., cosmopolitan, tropical and temperate) of species were taken from Subías (2004). Cosmopolitan distribution refers to taxa that occur in both temperate and tropical regions, rather than to those with global distribution. Tropical oribatid mites were collected in the Neotropic (Ecuador) but many also occur in the Palaeotropic. Similarly, the temperate species were mostly collected in the Palaearctic but many of them also occur in the Nearctic. Therefore, the taxon sampling was taken to represent high and low latitude species of oribatid mites.
Genomic DNA was extracted from single individuals using the DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany) with silica membrane columns and protease K from Genaxxon (Genaxxon BioScience, Ulm, Germany). Amplification of target genes was performed in 25 $icro$l volume. Primers for 18S rDNA were 5'-TACCTGGTTGATCCTGCCAG-3' (18Sforward) and 5'-AATGATCCTTCCGCAGGTTCAC-3' (18Sreverse) (Domes et al. 2007a). Primers for the D3-D5 region of 28S rDNA were 5'-GACCCGTCTTGAAGCACG-3' (28Sa) and 5'-CCCACAGCGCCAGTTCTGCTTACC-3' (28Sbout) (Tully et al. 2006). The 18S fragment was amplified at 57°C; the 28S fragment was amplified at 49°C using standard PCR protocols. PCR products were sequenced at Göttingen Genomics Laboratory (Institute of Microbiology and Genetics, Georg August University Göttingen, Germany), using the additional sequencing primers 18S554f 5'-AAGTCTGGTGCCAGCAGCCGC-3', 18S1282r 5'-TCACTCCACCAACTAAGAACGGC-3', 18S1150f 5'-ATTGACGGAAGGGCACCACCAG-3' and 18S614r 5'-TCCAACTACGAGCTTTTTAACC-3' (Domes et al. 2007a). For some species, PCR amplification of 18S rDNA was difficult and several shorter fragments were amplified using the above-mentioned primers and s6mod (Nikolaev et al. 2005) (modified for this study), 18S476fn (designed for this study) and Euk516f (Lara et al. 2008) in the combinations 18Sforward (5'-TACCTGGTTGATCCTGCCAG-3') and 18S614r, 18S554f and 18S1282r, s6mod (5'-CCGCGGTAATTCCAGCTC-3') and 18S1282r, 18S476fn (5'-GAGGTAGTGACGAAAAATAACAATACAGG-3') and 18S1282r, Euk516f (5'-GGAGGGCAAGTCTGGT-3') and 18S1282r, 18S1150f and 18Sreverse (5'-TAATGATCCTTCCGCAGGTTCAC-3'). Sequences KR081601-KR081690 were generated for this study; all other sequences were obtained from NCBI.
Phylogenetic analysis was based on a supermatrix of the ribosomal genes 18S rDNA (1,599-2,007 bp) and partial 28S rDNA (289-350 bp). Sequences were assembled in Sequencher 5.1 (Gene Codes Corporation, Ann Arbor, Michigan, USA); ambiguous positions were corrected using the chromatograms. Two preliminary alignments were generated in ClustalX v1.8 (Thompson et al. 1994) using default parameters for the 18S and the 28S rDNA sequences. Sequences of the alignments were cut to the shortest sequence downloaded from NCBI. Trimmed alignments of both genes were concatenated in a single data set and a new alignment was calculated using the multiple alignment parameters gap opening 20 and gap extension 0.1. To compensate for possible long-branch attraction, a NJ tree was calculated in SeaView v4.2.3 (Gouy et al. 2010) and used as guide tree for a new alignment in ClustalX with the same parameters. The final alignment had a length of 2,412 bp, consisting of 2,077 bp 18S and 335 bp 28S rDNA.
The model of sequence evolution was estimated in PartitionFinder v 1.1.1. (Lanfear et al. 2012). Phylogenetic trees were constructed using Bayesian Inference in MrBayes 3.2.6 (Huelsenbeck and Ronquist 2001) using JK as well as GTR+I+G and Maximum Likelihood in RAxML 7.0.4 (Stamatakis 2006) GTR+I+G. MrBayes was run for 5 million generations, reaching convergence with a standard deviation of split frequencies of 0.013630 (GTR + I + G) and 0.005798 (JK). RAxML was run with 1,000 bootstraps.
We used Mesquite 3.10 (Maddison & Maddison 2016) to map present day biogeographic distribution (temperate, tropical and cosmopolitan) as a character on the phylogenetic tree. Character history was traced using parsimony and likelihood to infer ancient distribution patterns. The Bayesian Inference tree (JK) provided the topology and the present day distribution of investigated species was coded as tropical, temperate or cosmopolitan. We also used the distribution extinction cladogenesis (DEC) biogeographical model (Ree & Smith 2008) for ancestral area reconstruction (implemented in BioGeoBEARS in R; Matzke 2014; R Core Team, 2014). The likelihood analysis in Mesquite (data not shown) and the DEC analysis (Fig. S3) showed a weaker resolution than the parsimony analysis; probably since we did not sample enough taxa covering the distribution of oribatid mites all over the world.
Topologies of the BI (JK; GTR + I + G) and ML trees were very similar (Fig. 1; Figs. S1; S2 in Electronic Appendix). Differences were restricted to the resolution of the backbone and node support of early-derived Enarthronota, Palaeosomata and Parhyposomata. Generally, the three trees did not contradict each other, but the ML tree had dichotomous splits with low bootstrap support (< 20) while the BI tree (GTR + I +G) had polytomies, especially in early-derivative groups (Fig. S1 in Electronic Appendix), whereas the BI tree (JK) was fully resolved with high support.
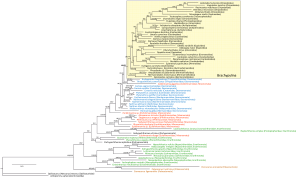

Tree topologies generally were conform to previously published phylogenies (Norton et al. 1993, Pachl et al. 2012). Brachypylina were most derived, Desmonomata and Mixonomata were intermediate and paraphyletic, and early-derivative oribatid mites (Enarthronota, Parhyposomata, Palaeosomata) were at the basis of the tree. Ameroidea, represented by three taxa, was not resolved as a monophyletic group, as one taxon (Haplobelba sp.) clustered together with Ceratozetidae, Phenopelopoidea and Oripodoidea. Among middle-derived oribatid mites (Desmonomata, Mixonomata), taxonomic groups were well resolved except for Collohmannia gigantea and Eulohmannia ribagai, two Mixonomata placed within Desmonomata, and among early-derivative oribatid mites. Among early-derivative oribatid mites (Enarthronota, Palaeosomata, Parhyposomata) only Palaeosomata formed a monophyletic group. Enarthronota formed two monophyletic clades, and one paraphyletic cluster comprising other Enarthronota, Mixonomata and Parhyposomata.
In the following we present the findings of the parsimony analysis in Mesquite (Fig. 2). Palaeosomata were represented by two temperate and one tropical species. The ancestral state for this group was temperate with one derived tropical taxon. Enarthronota formed one large cluster, including three cosmopolitan, four tropical and three temperate species. The ancestral state was cosmopolitan with derived tropical taxa in the Mesoplophoridae. Hypochthoniidae and Lohmanniidae formed a tropical group with temperate and cosmopolitan offshoots. The other Enarthronota clustered together with Eulohmannia ribagai (Mixonomata) and two Parhyposomata; taxa which proved problematic to resolve in previous studies (Pachl et al. 2012).
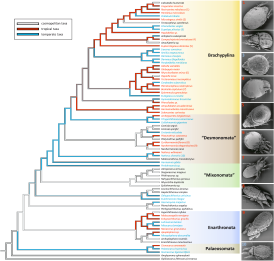

Mixonomata formed a large cosmopolitan clade. The three temperate members of this group were located on separate branches of the tree. In Desmonomata, the ancestral state of Nothridae was temperate with one derived tropical species. The ancestral state of Camisiidae was cosmopolitan; that of Trhypochthoniidae was temperate. Nanhermannioidea contained two tropical and one cosmopolitan species with a cosmopolitan ancestral state.
Brachypylina were the sister group to Trhypochthoniidae and Collohmannia sp., and their ancestral state was equivocal with an early-derived tropical taxon, Hermanielloidea. Ancestral state of derived Brachypylina (e.g., Achipteroidea, Oripodoidea) was tropical. Compared to early-derivative oribatid mites, Brachypylina had a reversed pattern with many tropical taxa and isolated, derived temperate or cosmopolitan offshoots. This pattern was prevalent in Hermannielloidea, Gymnodamaeoidea, Oppioidea and Carabodoidea. However, Damaeoidea and Gustavoidea within Brachypylina were temperate (Fig. 2).
Animal and plant diversity generally increases from polar to tropical regions. This pattern is also true for soil animals, e.g. oribatid mites, though less pronounced than in plants and aboveground invertebrates (Maraun et al. 2007, Decaens 2010). Using oribatid mites as a model taxon, we investigated if the tropics predominantly functioned as cradle or museum for soil animal species. We hypothesized that oribatid mites from the tropics are old and experienced less climatic change than those from biomes at higher latitudes, thereby functioning as museum of diversity. The results suggest that this hypothesis has to be rejected. As indicated by molecular phylogeny of temperate and tropical oribatid mite species tropical taxa more intensively diversified in 'recent' times (i.e. $im$ 250 mya, million years ago) as compared to temperate taxa (which evolved in Cambrian or even Precambrian times; Schaefer et al. 2010). This was especially true for the derived Brachypylina, supporting the view that radiation in the tropics surpasses that in temperate regions, and suggesting that the tropics function as cradle of oribatid mite diversity.
For understanding why the tropics function as cradle rather than museum of oribatid mite diversity the time scale of the radiations needs to be considered. Oribatid mites are an evolutionary very old group. Fossils are known from the Devonian (Norton et al. 1988), and trace fossils have been recovered from the Carboniferous (Labandeira et al. 1997); however, molecular clock estimations place the origin of oribatid mites into Cambrian or even Precambrian times (Schaefer et al. 2010). Therefore, early-derived lineages of oribatid mites experienced an exceptionally wide range of environmental conditions including those during the major mass extinction events. Further, irrespective of where they originated they were exposed to massive environmental changes due to plate tectonics. Certainly, a great number of lineages went extinct; however, a number of present day lineages must have survived all these changes.
Food relationships of early- and middle-derived taxa (Enarthronota, Desmonomata) indicates that ancient lineages of oribatid mites lived as decomposers or fungal feeders, contrasting the derived Brachypylina which include many species feeding on lichens and algae, and also predatory species (Maraun et al. 2011). Feeding on dead organic matter and association with thick organic layers is typical for oribatid mites of temperate and boreal regions where they reach maximum densities (Maraun & Scheu 2000). In contrast, in the tropics the density of oribatid mites is much lower than in high latitude forests suggesting that they suffer from fast decomposition of organic matter (Illig et al. 2010). High density of oribatid mites at high latitudes likely contributed to the survival of ancient lineages in temperate and boreal regions as the risk of extinction declines with population density, and the opposite likely is true for tropical regions. Present day occurrence of species of virtually all old lineages of oribatid mites in temperate biomes suggests that they survived in high latitude biomes and evolved there as detritivore animals. Many of the present day species of old lineages occurring in the tropics therefore may actually have colonized the tropics from high latitude biomes and thereby potentially widened their trophic niche.
Results of the present study suggest that the most diverse and derived lineage of oribatid mites, the Brachypylina, evolved in tropical regions. According to molecular clock dating this radiation started about 300 mya (Schaefer et al. 2010). This suggests that radiation started on the super continent Pangaea where oribatid mites were exposed mainly to tropical climate (Retallack & German-Heins 1994). Therefore, tropical regions of Pangaea may be viewed as the 'cradle' of diversity of Brachypylina. In fact, as documented by our analysis and proposed earlier (Maraun et al. 2008, de Moraes et al. 2011, Mumladze et al. 2015), Brachypylina dominate oribatid mite communities of extant tropical and subtropical regions. Compared to other taxa for which the tropics function as cradle of diversity, such as beetles (McKenna & Farrell 2006), the radiation of tropical oribatid mite species is much older and the tropics therefore may be viewed as an 'ancient cradle' of Brachypylina.
Our conclusion that early-derived lineages of oribatid mites evolved at conditions resembling those of temperate and boreal regions is supported by the fact that at times of their radiation tropical forests did not exist. Oribatid mites presumably evolved in the Cambrian or earlier (Schaefer et al. 2010), and warm forest-like vegetation resembling tropical forests of today only started to evolve about 360 mya. The evolution of tropical swamp forests, which covered Pangea around 300 mya (Retallack & German-Heins 1994), may then have triggered the radiation of tropical oribatid mites. Notably, early radiation of oribatid mite lineages in the Cambrian or earlier coincides with cold conditions after warming of snowball earth (Kirschvink 1992) suggesting that early-derived oribatid mite lineages in fact evolved at cold climatic conditions matching the fact that they flourish today in high latitude biomes.
Results of our study suggest that many of the early-derivative oribatid mite taxa currently have a cosmopolitan distribution. Similarly, Hammer & Wallwork (1979) suggested a cosmopolitan distribution of early- and middle-derived oribatid mite species supporting the view that oribatid mites are an old taxon that evolved prior to the split up of the today continents. In Cambrian times or earlier, oribatid mite species that colonized land likely were confronted with nutrient-poor habitats but, similar to today, they may have fed on organic material and fungi (Schaefer et al. 2010). Many oribatid mite species of those ancient taxa, such as Palaeosomata and Enarthronota, today still live as generalists in habitats such as sandy soils where they mainly consume fungi (Lehmitz & Maraun 2016). This narrow niche may have hampered radiation of these taxa indicating that resource-poor sandy soils act as a 'museum' for early-derivative oribatid mites.
The pattern that recently evolved oribatid mites such as Brachypylina dominate in the tropics, whereas more ancient taxa, such as Enarthronota, Desmonomata and Mixonomata flourish in high latitude forests is not without exceptions. A number of Brachypylina species occur in temperate forests (e.g., in our study Eupelops plicatus, Xenillus tegeocranus, Gymnodamaeus bicostatus) and many middle-derived oribatid mite taxa, such as Crotonia reticulata, Nanhermannia elegantissima and Lohmannia banksi, occur in tropical regions. These exceptions complicate general conclusions on the biogeographic origin of oribatid mites. However, they may be explained by ecological factors; e.g., Crotonia and Nanhermannia predominantly occur in microsites in the tropics, i.e. on the bark of trees (Crotonia) (Domes et al. 2007b) or at high altitude (Nanhermannia) (Maraun et al. 2013), resembling habitats of high latitude forests. Similarly, Brachypylina species of temperate forests, such as Eupelops plicatus and Gymnodamaeus bicostatus, also predominantly colonize trees (rather than soil). In fact, there is increasing evidence that at least in high latitude forests the bark of trees forms a unique habitat for oribatid mites with little overlap to species in soil (Skubala & Marzec 2013, Bluhm et al. 2015).
This study aimed at improving the understanding of processes responsible for the current diversity and distribution of oribatid mites. Oribatid mites are highly diverse and the taxon sampling of our study necessarily is limited. Therefore, the biogeographic inferences made in this paper need further proof based on a wider taxon sampling including species from other biogeographic regions.
Overall, oribatid mites likely originated in Cambrian or Precambrian times on a large ancient landmass as indicated by their present day cosmopolitan distribution. Those species probably were mainly decomposers and fungal feeders (as many of them still are today) adapted to cold climatic conditions. Parallel to the origin of tropical ecosystems around 300 mya and also parallel to the formation of Pangaea, oribatid mites (especially Brachypylina) radiated in tropical regions and split up into thousands of species. The tropics therefore might be viewed as "ancient cradle" of oribatid mite diversity. In contrast, cold habitats at high latitude where early-derived oribatid mites flourish might be viewed as "museum" of cold adapted early-derived oribatid mites which appear to have radiated much less than the (younger) tropical lineages.
Our findings, at least in part, also reject the "out of the tropics" hypothesis (origin and higher speciation but lower extinction rates in the tropics) since the tropics are evolutionary younger than oribatid mites which necessarily places the origin of oribatid mites in pre-tropical times. However, our data do not exclude the possibility that speciation rates in the tropics are higher in tropical oribatid mites than in oribatid mite taxa from temperate regions since species numbers of oribatid mites in the derived (and often tropical) Brachypylina are much higher than those of the early-derivative 'Macropylina'. Investigating speciation rates in oribatid mites remains a challenge for futures studies.
The paper benefitted from comments by Roy A. Norton, which we greatly appreciate. We thank three anonymous reviewers for their helpful comments. We thank Dorothea Hause-Reitner for help with the SEM pictures.
Supplementary materials are available online.
Abensperg-Traun M., Steven D. 1997 — Latitudinal gradients in the species richness of Australian termites (Isoptera) — Australian Journal of Ecology, 22: 471-476. doi:10.1111/j.1442-9993.1997.tb00698.x ![]()
Arita H.T., Vázquez-Domínguez E. 2008 — The tropics: cradle, museum or casino? A dynamic null model for latitudinal gradients of species diversity — Ecology Letters, 11: 653-663. doi:10.1111/j.1461-0248.2008.01197.x ![]()
Balogh J., Balogh P. 1988 — Oribatid mites of the neotropical region I. — In: Balogh J., Mahunka S. (eds.) The soil mites of the world. Akad. Kiadó, Budapest.
Balogh J., Balogh P. 1990 — Oribatid mites of the neotropical region II. — In: BaloghJ., Mahunka S. (eds.) The soil mites of the world. Elsevier, Amsterdam.
Balogh J., Balogh P. 2002 — Identification keys to the oribatid mites of the extra-holarctic regions. — Well-Press Publishing Limited, Miskolc.
Bluhm C., Scheu S., Maraun M. 2015 — Oribatid mite communities on the bark of dead wood vary with log type, surrounding forest and regional factors — Applied Soil Ecology, 89: 102-112. doi:10.1016/j.apsoil.2015.01.013 ![]()
Chown S.L., Gaston K.J. 2000 — Areas, cradles and museums: the latitudinal gradient in species richness — Trends in Ecology and Evolution, 15: 311-315. doi:10.1016/S0169-5347(00)01910-8 ![]()
Decaens T. 2010 — Macroecological patterns in soil communities — Global Ecology and Biogeography, 19: 287-302. doi:10.1111/j.1466-8238.2009.00517.x ![]()
de Moraes J., Franklin E., de Morais J.W., de Souza J.L.P. 2011 — Species diversity of edaphic mites (Acari: Oribatida) and effects of topography, soil properties and litter gradients on their qualitative and quantitative composition in 64 km2 of forest in Amazonia — Experimental and Applied Acarology, 55: 39-63. doi:10.1007/s10493-011-9451-7 ![]()
Domes K., Althammer M., Norton R.A., Scheu S., Maraun M. 2007a — The phylogenetic relationship between Astigmata and Oribatida as indicated by molecular markers. — Experimental and Applied Acarology, 42: 159-171. doi:10.1007/s10493-007-9088-8 ![]()
Domes K., Norton R.A., Maraun M., Scheu S. 2007b — Re-evolution of sex in oribatid mites breaks Dollo's law — Proceedings of the National Academy of Sciences of the USA, 104: 7139-7144. doi:10.1073/pnas.0700034104 ![]()
Erdmann G., Scheu S., Maraun M. 2012 — Regional factors rather than forest type drive the community structure of soil living oribatid mites (Acari, Oribatida) — Experimental and Applied Acarology, 57: 157-169. doi:10.1007/s10493-012-9546-9 ![]()
Gouy M., Guindon S., Gascuel O. 2010 — SeaView v4: a multiplatform graphical user interface for sequence alignment and phylogenetic tree building — Molecular Biology and Evolution, 27: 221-224. doi:10.1093/molbev/msp259 ![]()
Hammer M., Wallwork J.A. 1979 — Review of the world distribution of oribatid mites (Acari: Cryptostigmata) in relation to continental drift — Biologiske Skrifter, 22: 1-31.
Hillebrand H. 2004 — On the generality of the latitudinal diversity gradient — American Naturalist, 163: 192-211. doi:10.1086/381004 ![]()
Huelsenbeck J.P., Ronquist F. 2001 — MRBAYES: Bayesian inference of phylogenetic trees — Bioinformatics, 17: 754-755.
Illig J., Norton R.A., Scheu S., Maraun M. 2010 — Density and community structure of soil- and bark-dwelling microarthropods along an altitudinal gradient in a tropical montane rain forest — Experimental and Applied Acarology, 52: 49-62. doi:10.1007/s10493-010-9348-x ![]()
Jablonski D. 1993 — The tropics as a source of evolutionary novelty through geological time — Nature, 364: 142-144.
Jablonski D., Kaustuv R., Valentine J.W. 2006 — Out of the tropics: evolutionary dynamics of the latitudinal diversity gradient — Science, 314: 102-106.
Jablonski D., Belanger C.L., Berke S.K., Huang S., Krug A.Z., Roy K., Tomasovych A., Valentine J.W. 2013 — Out of the tropics, but how? Fossils, bridge species, and thermal ranges in the dynamics of the marine latitudinal diversity gradient — Proceedings of the National Academy of Sciences of the USA, 110: 10487-10494. doi:10.1073/pnas.1308997110 ![]()
Kempson D., Lloyd M., Ghelardi R.A. 1963 — A new extractor for woodland litter — Pedobiologia, 3: 1-21.
Kirschvink J. 1992 — Late proterozoic low-latitude global glaciation: the snowball Earth — In Schopf J.W., Klein C. (eds.) The proterozoic biosphere: a multidisciplinary study. Cambridge University Press, Cambridge.
Labandeira C.C., Phillips T.L., Norton R.A. 1997 — Oribatid mites and the decomposition of plant tissues in Paleozoic coal-swamp forests — Palaios, 12: 319-353.
Lanfear R., Calcott B., Ho S.Y.W., Guindon S. 2012 — PartitionFinder: combined selection of partitioning schemes and substitution models for phylogenetic analyses. — Molecular Biology and Evolution, 29: 1695-1701. doi:10.1093/molbev/mss020 ![]()
Lara E., Heger T.J., Ekelund F., Lamentowicz M., Mitchell E.A.D. 2008 — Ribosomal RNA genes challenge the monophyly of the Hyalospheniidae (Amoebozoa: Arcellinida) — Protist, 159: 165-176. doi:10.1016/j.protis.2007.09.003 ![]()
Lehmitz R., Maraun M. 2016 — Small-scale spatial heterogeneity of stable isotopes signatures ($elta$15N, $elta$13C) in Sphagnum sp. transfers to all trophic levels in oribatid mites — Soil Biology and Biochemistry, 100: 242-251. doi:10.1016/j.soilbio.2016.06.005 ![]()
Maddison W.P., Maddison D.R. 2016 — Mesquite: a modular system for evolutionary analysis. Version 3.10 — http://mesquiteproject.org
Mannion P.D., Upchurch P., Benson R.B.J, Goswami A. 2014 — The latitudinal biodiversity gradient through deep time — Trends in Ecology and Evolution, 29: 42-50. doi:10.1016/j.tree.2013.09.012 ![]()
Maraun M., Scheu S. 2000 — The structure of oribatid mite communities (Acari, Oribatida): patterns, mechanisms and implications for future research — Ecography, 23: 374-383.
Maraun M., Heethoff M., Schneider K., Scheu S., Weigmann G., Cianciolo J., Thomas R.H., Norton R.A. 2004 — Molecular phylogeny of oribatid mites (Oribatida, Acari): evidence for multiple radiations of parthenogenetic lineages — Experimental and Applied Acarology, 33: 183-201. doi:10.1023/B:APPA.0000032956.60108.6d ![]()
Maraun M., Schatz H., Scheu S. 2007 — Awesome or Ordinary? Global diversity patterns of oribatid mites — Ecography, 30: 209-216.
Maraun M., Illig J., Sandmann D., Krashevska V., Norton R.A., Scheu S. 2008 — Soil Fauna — In: Beck E., Benedix J., Kottke I. (eds.) Gradients in a tropical mountain ecosystem of Ecuador. Springer, Berlin.
Maraun M., Erdmann G., Fischer B.M., Pollierer M.M., Norton R.A., Schneider K., Scheu S. 2011 — Stable isotopes revisited: their use and limits for oribatid mite trophic ecology — Soil Biology and Biochemistry, 43: 877-882. doi:10.1016/j.soilbio.2011.01.003 ![]()
Maraun M., Fronczek S., Marian F., Sandmann D., Scheu S. 2013 — More sex at higher altitudes: changes in the frequency of parthenogenesis in oribatid mites in tropical montane rain forests — Pedobiologia, 56: 185-190.
Matzke N.J. 2014 — Package BioGeoBEARS: BioGeography with Bayesian (and likelihood) evolutionary analysis in R scripts. Version 0.2.1. 360 pp. — ( http://cran.r-project.org/web/packages/BioGeoBEARS/BioGeoBEARS.pdf ![]() ).
).
McKenna D., Farrell B.D. 2006 — Tropical forests are both evolutionary cradles and museums of leaf beetle diversity — Proceedings of the National Academy of Sciences of the USA, 103: 10947-10951. doi:10.1073/pnas.0602712103 ![]()
Miller K.G., Kominz M.A., Browning J.V., Wright J.D., Mountain G.S., Katz M.E., Sugarman P.J., Cramer B.S., Christie-Blick N., Pekar S.F. 2005 — The phanerozoic record of global sea-level change. Science, 310: 1293-1298. doi:10.1126/science.1116412 ![]()
Mittelbach G.G., Schemske D.W., Cornell H.V., Allen A.P., Brown J.M., Bush M.B., Harrison S.P., Hurlbert A.H., Knowlton N., Lessios H.A., McCain C.M., McCune A.R., McDade L.A., McPeek M.A., Near T.J., Price T.D., Ricklefs R.E., Roy K., Sax D.F., Schluter D., Sobel J.M., Turelli M. 2007 — Evolution and the latitudinal diversity gradient: speciation, extinction and biogeography — Ecology Letters, 10: 315-331. doi:10.1111/j.1461-0248.2007.01020.x ![]()
Moreau C.S., Bell C.D. 2013 — Testing the museum versus cradle tropical biological diversity hypothesis: phylogeny, diversification, and ancestral biogeographic range evolution of the ants — Evolution, 67: 2240-2257.
Mumladze L., Murvanidze M., Maraun M., Salakaia M. 2015 — Oribatid mite communities along an elevational gradient in Sairme gorge (Caucasus) — Experimental and Applied Acarology, 66: 41-51. doi:10.1007/s10493-015-9893-4 ![]()
Nikolaev S.I., Mitchell E.A.D., Petrov N.B., Berney C., Fahrni J., Pawlowski J. 2005 — The testate lobose amoebae (Order Arcellinida Kent, 1880) finally find their home within Amoebozoa — Protist, 156: 191-202.
Norton R.A., Behan-Pelletier V.M. 2009 — Suborder Oribatida — In: Krantz G.W., Walter D.E. (eds.) A Manual of Acarology. Texas Tech University Press, Texas.
Norton R.A., Bonamo P.M., Grierson J.D., Shear W.A. 1988 — Oribatid mite fossils from a terrestrial Devonian deposit near Gilboa, New York — Journal of Paleontology, 62: 259-269. doi:10.1017/S0022336000029905 ![]()
Norton R.A., Kethley J.B., Johnston D.E., O'Connor B.M. 1993 — Phylogenetic perspectives on genetic systems and reproductive modes of mites — In: Wrensch D.L., Ebbert M.A. (eds.) Evolution and Diversity of Sex Ratios. Chapman and Hall, New York. doi:10.1007/978-1-4684-1402-8$_$2 ![]()
Pachl P., Domes K., Schulz G., Norton R.A., Scheu S., Schaefer I., Maraun M. 2012 — Convergent evolution of defense mechanisms in oribatid mites (Acari, Oribatida) shows no "ghosts of predation past" — Molecular Phylogenetics and Evolution, 65: 412-420. doi:10.1016/j.ympev.2012.06.030 ![]()
Proches S. 2001 — Back to the sea: secondary marine organisms from a biogeographical perspective — Biological Journal of the Linnean Society, 74: 197-203. doi:10.1111/j.1095-8312.2001.tb01386.x ![]()
Ree R.H., Smith S.A. 2008 — Maximum likelihood inference of geographic range evolution by dispersal, local extinction, and cladogenesis — Systematic Biology, 57: 4-14. doi:10.1080/10635150701883881 ![]()
Retallack G.J., German-Heins J. 1994 — Evidence from paleosols for the geological antiquity of rain forest — Science, 265: 499-502.
Rohde K. 1992 — Latitudinal gradients in species diversity – the search for the primary cause — Oikos, 65: 514-527.
Rolland J., Condamine F.L., Beeravolu C.R., Jiguet F., Morlon H. 2015 — Dispersal is a major driver of the latitudinal diversity gradient of Carnivora — Global Ecology and Biogeography, 24: 1059-1071. doi:10.1111/geb.12354 ![]()
Roy K., Jablonski D., Valentine J.W., Rosenberg G. 1998 — Marine latitudinal diversity gradients: Tests of causal hypotheses — Proceedings of the National Academy of Sciences of the USA, 95: 3699-3702. doi:10.1073/pnas.95.7.3699 ![]()
Runkel A.C., Mackey T.J., Cowan C.A., Fox, D.L. 2010 — Tropical shoreline ice in the late Cambrian: Implications for Earth's climate between the Cambrian Explosion and the Great Ordovician Biodiversification Event — GSA Today, 20: 4-10. doi:10.1130/GSATG84A.1 ![]()
Schaefer I., Norton R.A., Scheu S., Maraun M. 2010 — Arthropod colonization of land – linking molecules and fossils in oribatid mites (Acari, Oribatida) — Molecular Phylogenetics and Evolution, 57: 113-121. doi:10.1016/j.ympev.2010.04.015 ![]()
Skubala P., Marzec A. 2013 — Importance of different types of beech dead wood for soil microarthropod fauna — Polish Journal of Ecology, 61: 545-560.
Stamatakis A. 2006 — RAxML-VI-HPC: Maximum Likelihood-based phylogenetic analyses with thousands of taxa and mixed models — Bioinformatics, 22: 2688-2690.
Stebbins G.L. 1974 — Flowering Plants. Evolution above the species level — Belknap Press.
Stenseth N.C. 1984 — The tropics: cradle or museum? — Oikos, 43: 417-420. doi:10.2307/3544168 ![]()
Subías L.S. 2004 — Listado sistematico, sinonimico y biogeografico de los acaros oribatidos (Acariformes, Oribatida) del Mundo (1758-2002) — Graellsia, 60: 3-305. (updated in February 2016; ( http://escalera.bio.ucm.es/usuarios/bba/cont/docs/RO$_$1.pdf ![]() ))
))
Subías L.S., Arillo A. 2002 — Oribatid mite fossils from the upper Devonian of South Mountain, New York and the lower Carboniferous of County Antrim, Northern Ireland (Acariformes, Oribatida) — Estacion Museum de Ciencia Natura de Álava, 17: 93-106.
Thompson J.D., Higgins D.G., Gibson T.J. 1994 — CLUSTAL W: improving the sensitivity of progressive multiple alignment through sequence weighting, position specific gap penalties and weight matrix choice — Nucleic Acids Research, 22: 4673-4680. doi:10.1093/nar/22.22.4673 ![]()
Tully T., D'Haese C.A., Richard M., Ferriere R. 2006 — Two major evolutionary lineages revealed by molecular phylogeny in the parthenogenetic Collembola species Folsomia candida — Pedobiologia, 50: 95-104.
Walter D.E., Proctor H.E. 2013 — Mites: Ecology, evolution and behavior. Life at a microscale — Springer, Dordrecht.
Weigmann G. 2006 — Hornmilben (Oribatida). Die Tierwelt Deutschlands, 76. Teil — Goecke and Evers, Keltern.
Wiens J.J., Graham C.H., Moen D.S., Smith S.A., Reeder T.W. 2006 — Evolutionary and ecological causes of the latitudinal diversity gradient in hylid frogs: treefrog trees unearth the roots of high tropical diversity — American Naturalist, 168: 579-596. doi:10.1086/507882 ![]()
Willig M.R., Kaufman D.M., Stevens R.D. 2003 — Latitudinal gradients of Biodiversity: Pattern, process, scale, and synthesis — Annual Reviews of Ecology and Systematics, 34: 273-309. doi:10.1146/annurev.ecolsys.34.012103.144032 ![]()
acarologia_4148_Electronic_Appendix_S1_Bayes_GTR.pdf
acarologia_4148_Electronic_Appendix_S2_RAxML.pdf
acarologia_4148_Electronic_Appendix_S3_DEC_model.pdf



2016-04-21
Date accepted:
2016-09-19
Date published:
2016-12-20
Edited by:
Migeon, Alain

This work is licensed under a Creative Commons Attribution 4.0 International License
2017 Pachl, Patricia; Lindl, Ann Christin; Krause, Alena; Scheu, Stefan; Schaefer, Ina and Maraun, Mark
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)