




Sublethal effects of spiromesifen on life table traits of Tetranychus urticae (Acari: Tetranychidae) and Neoseiulus californicus (Acari: Phytoseiidae)
Rajaee, Fahimeh1
; Ghane-Jahromi, Mojtaba2
; Maroofpour, Nariman  3
and Sedaratian-Jahromi, Amin4
3
and Sedaratian-Jahromi, Amin4
1Department of Plant Protection, Faculty of Agriculture, Yasouj University, Yasouj, Iran.
2Department of Plant Protection, Faculty of Agriculture, Yasouj University, Yasouj, Iran.
3✉ Department of Plant Protection, Faculty of Agriculture, University of Tabriz, Tabriz, Iran.
4Department of Plant Protection, Faculty of Agriculture, Yasouj University, Yasouj, Iran.
2022 - Volume: 62 Issue: 3 pages: 772-785
https://doi.org/10.24349/uja8-5ks2Original research
Keywords
Abstract
Introduction
The two-spotted spider mite, Tetranychus urticae Koch (Acari: Tetranychidae), is one of the most well-known and economically important pests with a broad host plant range in greenhouse and field conditions. Reduced plant vigor, prevention of photosynthesis, webbing, fine stippling, leaf yellowing, leaf drop, and even plant death result from the feeding and sucking of this pest and finally causes considerable yield losses (Li et al. 2017; Susurluk and Gürkan 2020; Shang et al. 2022).
The biological control by using Phytoseiidae mites is successful in greenhouse conditions (Kang et al. 2018; Kumari et al. 2017). One of the primary natural enemies used against this pest is Neoseiulus californicus (McGregor) (Acari: Phytoseiidae) (Rhodes et al. 2006). This natural enemy is a predator of tetranychid mites, particularly T. urticae (McMurtry et al. 2013). This species is used against T. urticae around the world (Sarbaz et al. 2017). Nowadays, the integrated pest management strategy uses biological and other controls in complementary ways (Dara 2017). On the other hand, the first way to control this pest is to apply chemical pesticides (Asadi et al. 2019). Systemic compounds like spiromesifen from the tetronic acid derivatives group are developed to control this species. Spiromesifen as an insecticidal/acaricidal with a specific mode of action is utilized in farms and greenhouses (Asadi et al. 2019; Sarbaz et al. 2017). This compound acts as a lipid biosynthesis inhibitor (Kumari et al. 2017).
Because of several abiotic factors, target and non-target populations receive sublethal concentrations of insecticides in field crops (Dai et al. 2021; Guedes et al. 2016). The sublethal concentrations can cause impairments in physiological and behavioral attributes (Desneux et al. 2007). Meanwhile, the predators can be exposed to pesticides by different approaches like direct contact with spray droplets, consuming the contaminated diet when feeding on food (e.g., pollen, prey), and foliar residues when exploring the crop (Dai et al. 2021; Lu et al. 2012). Therefore, the assessment of lethal and sublethal effects on these populations are important to be performed (Castro et al. 2012; De Castro et al. 2013; Maroofpour et al. 2021). The life table response experiments are more helpful in predicting the impacts of acaricides on the population of target and non-target species. These experiments reveal the overall impacts and the changes in the population growth rate (Sangak Sani et al. 2019). Moreover, the accurate baseline data on the susceptibility of target mite species to acaricides is one of the essential factors in managing acaricides (Kumari et al. 2017). There are several studies about the short-term effects of spiromesifen on T. urticae and N. californicus, whereas the potential long-term influence of this compound has been scarcely investigated (Kumari et al. 2017; Phukan et al. 2017; Sarbaz et al. 2017; Mollaloo et al. 2017; Marcic et al. 2010). Therefore, this study aimed to assess these general effects in successive generations for T. urticae and N. californicus exposed to a sublethal dose of spiromesifen.
Material and methods
Insecticide
Commercial formulations of spiromesifen (Oberon® SC, 240 g a.i/L), was purchased from Giah Bazr Alvand Co., Iran.
Biological materials
The original colony of T. urticae was obtained from infested cucumber plants collected in cucumber fields at the Yasuj region (Iran). Two-spotted spider mite was continuously reared on cucumber plants (Cucumis sativus L. var. Emperator) in the laboratory at 25 ± 2 °C of temperature, 65 ± 5% relative humidity (RH), and a photoperiod of 16: 8 h (L: D). Cucumber plants were grown in plastic pots (18 cm in diameter and 19 cm high), and at the seven-leaf stage were used for two-spotted spider mite rearing and all the experiments. The plastic pots were filled by coco peat and perlite (1:1). The mite-infected plants were weekly replaced with new and uncontaminated plants.
The colony of N. californicus was purchased from Gyah (Karaj, Iran). Individuals of N. californicus were reared on two-spotted spider mite-infested leaves of cucumber plants at 25 ± 2 °C of temperature, 65 ± 5% relative humidity (RH), and a photoperiod of 16: 8 h (L: D). The Petri dishes contain cucumber leaf discs (5 cm in diameter) and soaked cotton. New Petri dishes were supplemented at regular intervals (three days) to maintain the culture for experimentation.
Lethal toxicity on the two-spotted spider mite
The concentration-mortality bioassay was conducted on protonymphs of two-spotted spider mite. Adult females were selected from the population and placed on Petri dishes to establish a synchronous mite culture. After 12 h, the obtained protonymphs were transferred to new Petri dishes to assess the sublethal effects. Six serial concentrations (12, 60, 120, 180, 204 and 240 g a.i/L) of the insecticide were sprayed on the Petri dishes (6 cm in diameter) containing twenty similar-age protonymphs (\textless12 h old) by using a Potter tower (Burkard Scientific, Uxbridge, UK). The Petri dishes contain cucumber leaf discs (5 cm in diameter) and soaked cotton. Distilled water was used for the control group. The Petri dishes were kept in the growth chamber maintained at 25 ± 2 °C of temperature, 65 ± 5% relative humidity (RH), and a photoperiod of 16:8h (L:D). The two-spotted spider mite mortality was recorded after 48 h, and the experiments were replicated three times.
Sublethal effect on the first generation of the two-spotted spider mite
In this experiment, the sublethal impact of spiromesifen at low concentration (LC20) estimated for the protonymphs as described above was carried out on 200 protonymphs (\textless12 h old) of the two-spotted spider mite. The protonymphs were sprayed with 3 ml of LC20 concentration and distilled water (control treatment) following the methods as mentioned earlier. After 24 h of treatment, 100 surviving protonymphs were transferred to individual Petri dishes containing cucumber leaf discs (5 cm in diameter) along with soaked cotton. The surviving protonymphs were kept there until adulthood. The Petri dishes were observed daily for recorded life table traits (e.g., survival, mortality, and progeny production).
Transgenerational effects on life table traits of the two-spotted spider mite
The fresh eggs (≤12 h) were laid by T. urticae females exposed to the low concentration (LC20). One hundred eggs were transferred to individual Petri dishes containing cucumber leaf discs (5 cm in diameter) along with soaked cotton and maintained until the adult phase at the same conditions as the other experiments. The Petri dishes were checked daily to record life table traits (e.g., survival, mortality, and progeny production).
Sublethal effects of spiromesifen on the parental generation of Neoseiulus californicus
The sublethal evaluation was carried out on 90 similar-aged female mites (< 24 h old). For this purpose, 3 ml of LC20 concentration and distilled water (control treatment) were sprayed on the leaf discs (5 cm in diameter) using a Potter tower. After 24 h exposure, similar-aged female mites were transferred to the treated leaf discs. Sixty surviving females were removed and transferred to separate clean Petri dishes after 48 h of treatment. Then, females were paired with males and fed with ten protonymphs daily. Females were observed daily until death to calculate fecundity and survival. All experiments were conducted under controlled conditions in a growth chamber at 25 ± 2 °C, 60 – 70% RH, and a photoperiod of 16:8h (L:D).
Transgenerational effects and consumption rate of F1 generation of Neoseiulus californicus
This investigation was carried out with the fresh eggs obtained from the previous experiment parental. Sixty similar-aged eggs (\textless12 h old) were transferred to separate Petri dishes and monitored daily until the last mite died. The Petri dishes contain cucumber leaf discs (5 cm in diameter) and soaked cotton. After the emergence of adults, their females and males were paired, and ten protonymphs were daily offered to the predatory mites, and the number of prey mites consumed was recorded. The Petri dishes were kept under controlled conditions in a growth chamber at 25 ± 2 °C, 60 – 70% RH, and a photoperiod of 16:8h (L:D).
Statistical analyses
The data of concentration-response bioassay were subjected to probit analysis using the SPSS software (SPSS 2011), after using Abbott's formula for correction of natural mortality (Abbott 1925). The time-mortality were determined based on Cox regression, with right-censored data and surviving time as independent variables (SPSS, 2011).
The computer program TWOSEX-MS Chart, based on the age–stage, two-sex life-table theory was used to analyze the life-table and population parameters for both mite species (Chi 1988; Chi 2020b; Chi and Su 2006; Chi et al. 2020; Tuan et al. 2014). The bootstrap technique calculated the standard errors of all population parameters with 100,000 replicates (Chi 2020b).
The CONSUME-MS Chart analyzed daily consumption rates, and the standard errors were calculated through the bootstrap technique with 100,000 replicates (Chi 2020a; Chi and Yang 2003). The consumption parameters estimated were age-specific net consumption rate (qx ), net consumption rate (C0), finite predation rate (ω), stable consumption rate (ψ), age-specific consumption rate (kx ), and transformation rate (Qp ).
Results
Spiromesifen toxicity against Tetranychus urticae
The relative toxicity of spiromesifen to the two-spotted spider mite is shown in Table 1. According to the results, the values 41.01, 133.4, and 803.1 g a.i/L were estimated for LC20, LC50, and LC90, respectively.


Sublethal effect on the first generation of the two-spotted spider mite
Sublethal dose of spiromesifen had a negative effect on the development of treated protonymphs (Table 2). The significant effects of the low concentration were observed for most parameters except the adult preovipositional period (F= 138.18; P = 0.104) and the total preovipositional period (F= 35.45; P = 0.410). Larva-deutonymph (F= 1232.41; P \textless0.001) and preadult parameters (F= 274.6; P = 0.046) significantly increased with the insecticide. Meanwhile, total longevity (F= 1142.18; P \textless0.001), oviposition days (F= 9870.32; P \textless0.001), fecundity (F= 12093.68; P \textless0.001), the longevity of females (F= 3768.33; P \textless0.001) and males (F= 286.09; P \textless0.001) showed a significant decrease due to the treatments compared to the control.

Besides, significant differences were also observed in the intrinsic rate of increase (r) (F= 2433.78; P \textless0.001), net reproductive rate (R0) (F= 3850.44; P \textless0.001), and generation time (T) (F= 1218.07; P \textless0.001) (Table 3). There was no significant effect of spiromesifen in the age-stage survival rate (sxj ) of each age-stage (Fig. 1). However, the LC20 concentration indicated adverse effects on the age-specific survival rate (lx ), the age-specific fecundity (mx ) and maternity (lxmx ) (Fig. 2). The egg-laying starting age was delayed, and the peak of fecundity was 5 and 1.8 eggs/female at the control, and LC20, respectively. Meanwhile, the life expectancy (exj ) (Fig. 3) and the age-stage-specific reproductive values (vxj ) (Fig. 4) were decreased after treatment.





Transgenerational effects on the two-spotted spider mite
Effects of sublethal dose of spiromesifen on the two-spotted spider mite are presented in Table 2. Based on the results, the treatment of F0 generation by the low concentration of spiromesifen had no significant negative effect on Larva-deutonymph (F= 18.99; P = 0.605) and preadult parameters (F= 123.76; P = 0.188) of the F1 generation of T. urticae. In contrast, total longevity (F= 906.09; P \textless0.001), the adult preovipositional period (F= 285.27; P \textless0.001), oviposition days (F= 2512.95; P \textless0.001), and fecundity (F= 2369.37; P \textless0.001) were significantly decreased (Table 2). Furthermore, the intrinsic rate of increase (r) (F= 1203.15; P \textless0.001), net reproductive rate (R0) (F= 2764.73; P \textless0.001), and generation time (T) (F= 527.46; P = 0.015) of the F1 generation were significantly decreased (Table 3).
The negative effects were observed in the age-stage survival rate (sxj ) (Fig. 1). The low insecticide concentration decreased the survival rate and adult duration in both sexes of T. urticae. Moreover, the LC20 had negative effects on the age-specific survival rate (lx ), the age-specific fecundity (mx ), and maternity (lxmx ) (Fig. 2). The egg-laying starting age was delayed, and the peak of fecundity was 3.4 and 1.7 eggs/female at the control, and LC20, respectively. Meanwhile, the life expectancy (exj ) (Fig. 3) and the age-stage-specific reproductive values (vxj ) (Fig. 4) were decreased after treatment with the low insecticide concentration.
Sublethal effects on the parental generation of Neoseiulus californicus
The sublethal effects on the F0 generation of N. californicus are reported in Table 4. The longevity of female adults (F= 12712.18; P \textless0.001), fecundity (F= 9259.92; P \textless0.001), and oviposition days (F= 15388.94; P \textless0.001) significantly decreased after exposure. Meanwhile, the net reproductive rate (F= 9259.92; P \textless0.001), and generation time (F= 1960.70; P \textless0.001) was significantly decreased except the intrinsic rate of increase (F= 184.04; P= 0.062). Based on the results, the start of egg-laying was delayed in the low insecticide concentration. On the other hand, the highest age-specific fecundities were decreased in the LC20 (1.7 eggs/female) compared to the control (3.4 eggs/female) (Fig. 5).


Transgenerational effects and consumption rate of F1 generation of Neoseiulus californicus
Spiromesifen had negative effects on the life-history traits in F1 from F0 treated generation of N. californicus (Table 5). The significant decrease registered for larva-deutonymph (F= 212.73; P = 0.037), preadult (F= 569.67; P \textless0.001), adult preovipositional period (F= 206.13; P = 0.015), oviposition days (F= 566.17; P \textless0.001), and fecundity (F= 1598.24; P \textless0.001). Whereas there was no significant difference in longevity of egg (F= 198.74; P = 0.051), total longevity (F= 4.18; P = 0.77), total preovipositional period (F= 31.71; P = 0.34), and longevity of female (F= 1.75; P = 0.82) and male (F= 1.07; P = 0.77). Moreover, the LC20 significantly decreased the intrinsic rate of increase (F= 319.83; P = 0.015) and generation time (F= 729.20; P \textless0.001) (Table 6).

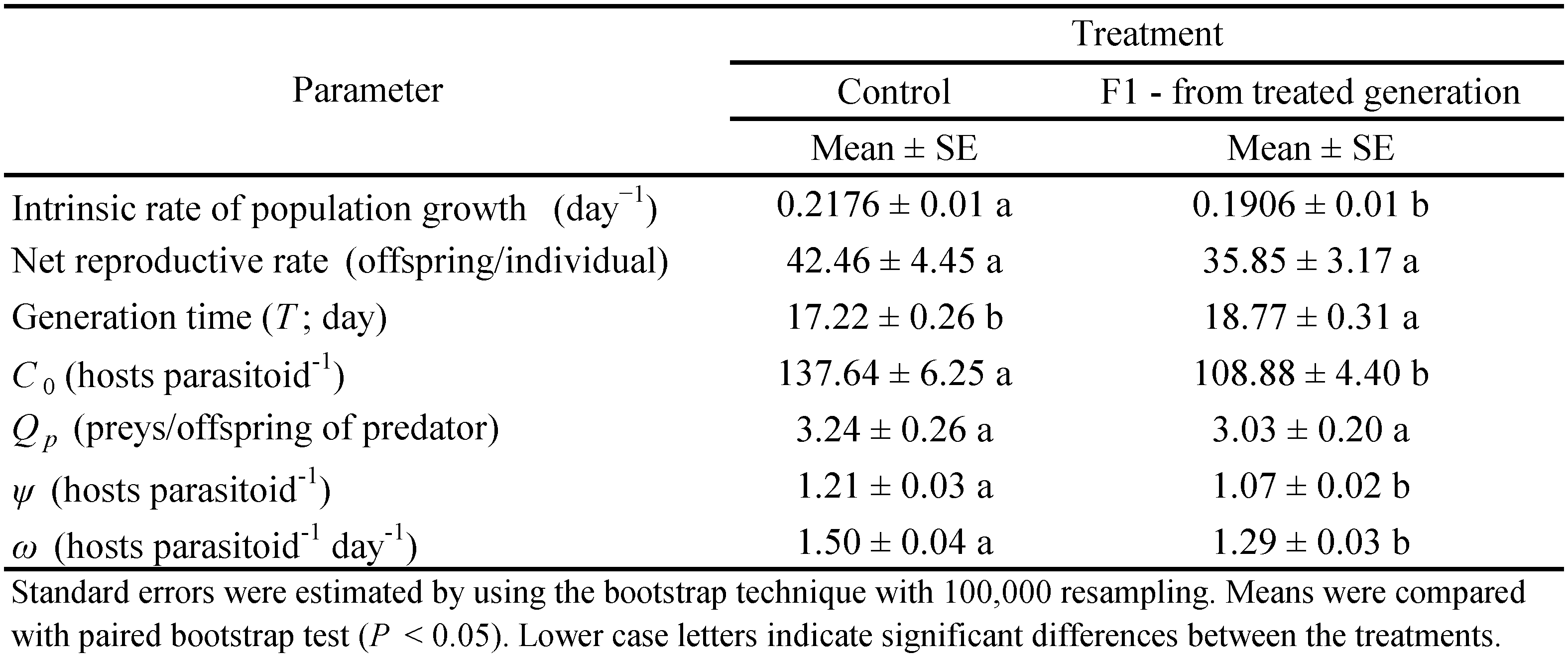
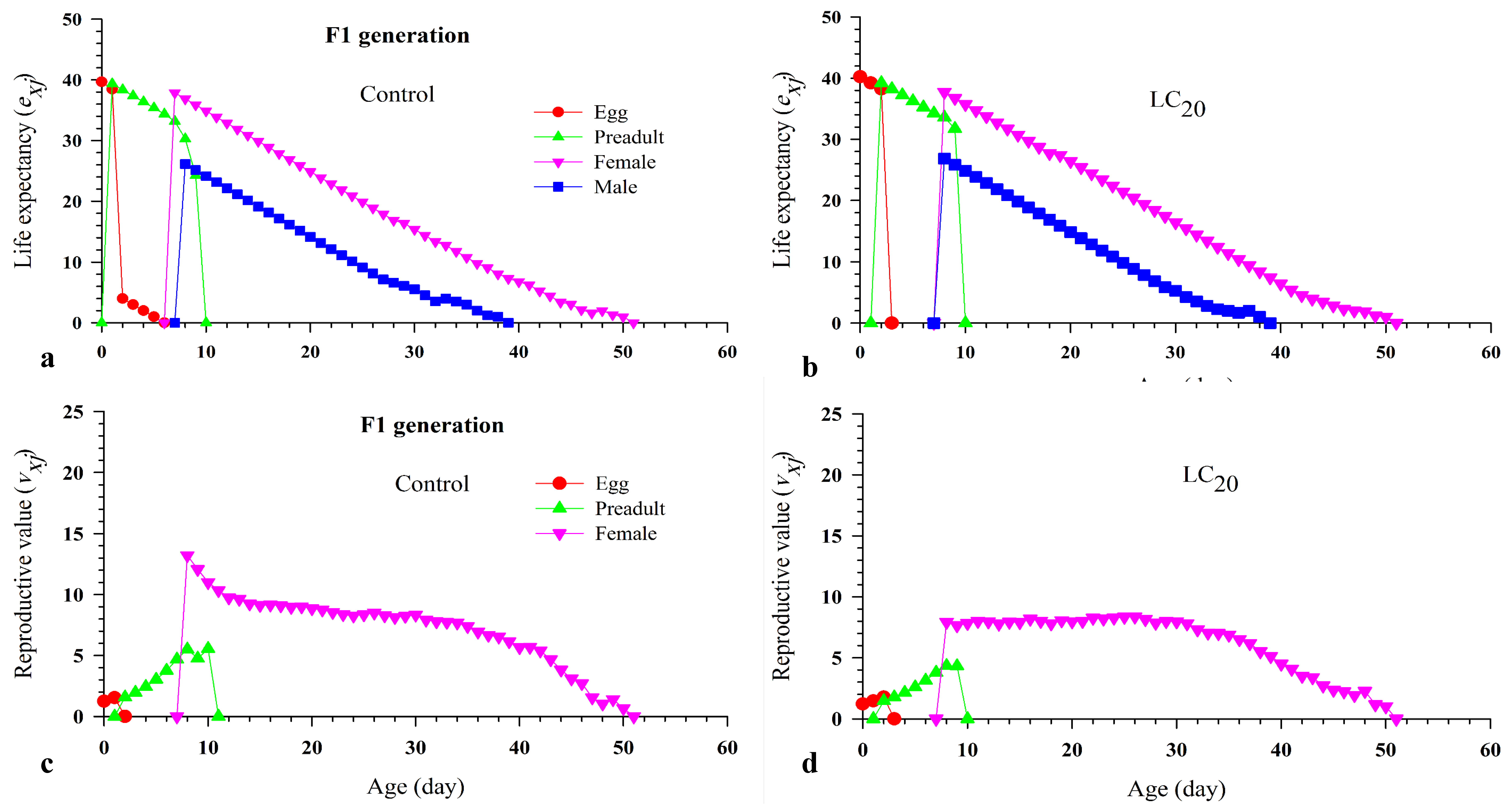
Accordingly to results, there was no negative effect on the age-stage survival rate (sxj ) of each mite age-stage and the age-specific survival rate (lx), the age-specific fecundity (mx), and maternity (lxmx) (Fig. 1 and 2). Furthermore, the LC20 had no negative effect on the life expectancy (exj ) (Fig. 6) and the age-stage-specific reproductive values (vxj ) (Fig. 7).

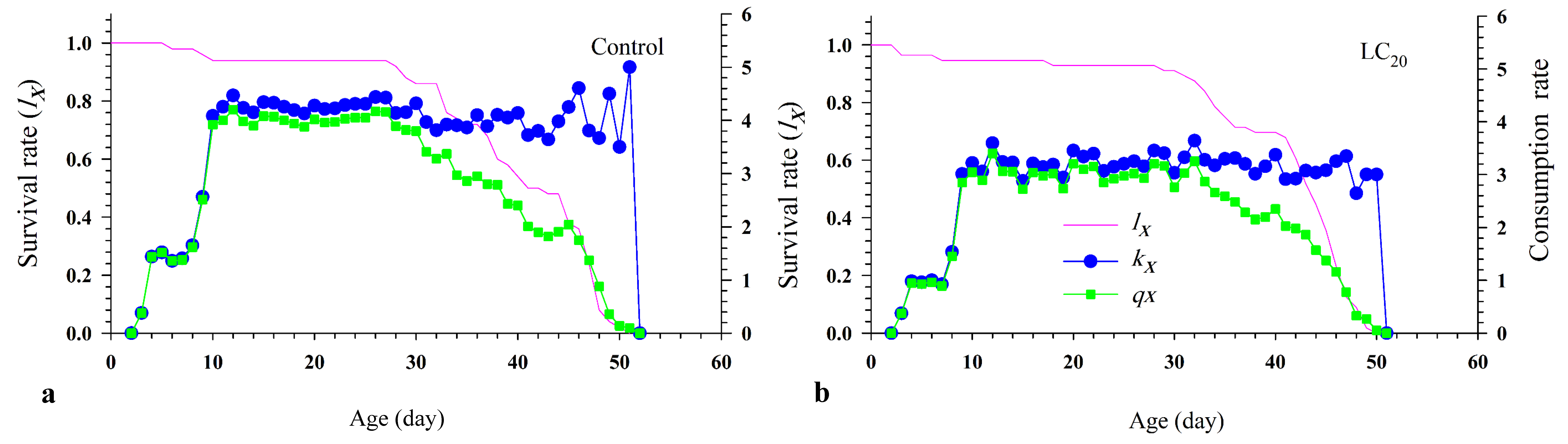
The sublethal effects on the consumption rate of N. californicus are summarized in Table 6. The low insecticide concentration significantly decreased net consumption rate (C0) (F= 764.97; P \textless0.001), finite predation rate (ω) (F= 751.99; P \textless0.001), stable consumption rate (ψ) (F= 668.86; P \textless0.001), except transformation rate (Qp ) (F= 22.83; P = 0.49) (Table 6). Subsequently, The LC20 decreased the age-specific parasitism rate (kx ) and age-specific net parasitism rate (qx ).
Discussion
Towards the rational application of insecticides, investigation on the effects of insecticides is an essential issue because insect populations are frequently exposed to lethal or sublethal concentrations of insecticides in the field (Guedes et al. 2016; Majidpour et al. 2020). Limited studies have been conducted separately on the effects of spiromesifen on T. urticae and their natural enemies, most of which have been on the first generation of insects (Kumari et al. 2017; Marcic et al. 2010, 2009; Mollaloo et al. 2017; Sarbaz et al. 2017). A comprehensive evaluation of the total impact on insect populations is essential to include the short-term effects of insecticides and the impact on the next generation, which can have important implications for the success of an IPM program (Müller 2018; Ali et al. 2017). Our study has clarified the sublethal effects of pesticides on both pests and their natural enemies in successive generations, how residual pesticides affect natural enemies, and how natural enemies respond to environmental xenobiotics. Based on the results, spiromesifen had a significant negative effect on most life table parameters of the pest population. These results showed this pesticide had suitable control in the successive generation of T. urticae. Besides, the LC20 concentration had a significant negative effect on most life table parameters of N. californicus in the successive generation.
Consistent with the present study, spiromesifen showed high toxicity in the female stages of the two-spotted spider mite in one-third of the recommended concentration (Kumari et al. 2017). In another study, spirodiclofen had a significant negative impact on most parameters (development time, longevity and fecundity) of T. urticae in the different sublethal concentrations (LC5, LC15 and LC35) (Sangak Sani et al. 2019). The obtained results of the present study are in the same line as those reported by Kumari et al. (2017) and Sangak Sani et al. (2019). Similarly, Phukan et al. (2017) reported that spiromesifen in the recommended concentration cause reduced the fecundity potential in females and the mite population of T. urticae decreased consequently. Meanwhile, (Marcic et al. 2010) showed that spiromesifen at different concentrations had a significant negative impact on the fecundity and net fertility of T. urticae. In another study, Marcic et al. (2009) recorded that the two-spotted spider mite population growth rates and fecundity were significantly reduced after exposure with spiromesifen. Besides, Marcic (2007) reported that spirodiclofen significantly affected the survival rate and total fecundity of T. urticae. Consistent with the present study, some investigations demonstrated that spiromesifen have toxicity and pronounced residual effects on the fecundity and development stages of T. urticae (Dekeyser 2005; Nauen and Schnorbach 2005; Sato et al. 2011; Wachendorff et al. 2002). Overall, the previous assessments only showed adverse effects of spiromesifen in the first generation on T. urticae, but the present study demonstrated that this compound has appropriate long-term effects for influencing the pest population.
On the other hand, the effects of spiromesifen have been studied in different natural enemies. A study by Sarbaz et al. (2017) demonstrated that spiromesifen and spirodiclofen at different concentrations decreased the longevity of N. californicus. They also reported that spiromesifen and spirodiclofen affected the intrinsic rate of increase (r) and net reproductive rate (R0), and these parameters decreased after exposure. In another study, the low concentrations (LC5, LC10, and LC15) of spiromesifen significantly decreased the longevity and population parameters (except mean generation time (T)) of N. californicus. However, this compound had no significant negative effects on the development stage of this predator (Mollaloo et al. 2017). In consistence with the present study, Lee and Kim (2015) found that spirodiclofen, spiromesifen did not affect the egg stage of N. californicus. Meanwhile, low toxicity was recorded for the nymphs, adult females, and reproduction. The reduced food uptake due to acaricide effects might partially explain these reductions (Hamedi et al. 2009). Our results are in line with Cloyd et al. (2006), who reported that spiromesifen decreased the longevity in N. californicus. Moreover, reduction of the intrinsic rate of population growth (r) in females of N. californicus was recorded when this predator was exposed to spirodiclofen and fenazaquin (Maroufpoor et al. 2016). However, the low concentrations of spirodiclofen (LC10, LC20, and LC30) had no significant effect on population parameters of offspring Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae) (Alinejad et al. 2016). This contrast may be due to species differences, different susceptibility of phytoseiid species, experimental conditions, and tested acaricides.
Conclusion
According to the results, spiromesifen is highly toxic to the two-spotted mite, even at low concentration. On the other hand, the adverse effects of this insecticide affected the subsequent generation. Meanwhile, this compound negatively affected the different generations of N. californicus. Overall, the obtained results of this research under laboratory conditions emphasized the significance of evaluating sublethal effects of spiromesifen on the two-spotted mite and its predator N. californicus and determining how these effects may be interpreted to population dynamics in the field. Further research into spiromisifin's impact on target and non-target organisms under semi-field and field conditions is vital to improving our understanding of pest management by considering the interaction of chemical and biological control.
Conflict of interest
The authors declare that they have no conflict of interest.
References
- Abbott W.S. 1925. A method of computing the effectiveness of an insecticide. J. Econ. Entomol., 18: 265-267 https://doi.org/10.1093/jee/18.2.265a
- Ali S.S., Ahmed S.S., Rizwana H., Bhatti F., Khoso A., Mengal M.I., Jatoi J.H., Bugti A., Rind M.A., Shahwani S.A. 2017. Efficacy of different bio-pesticides against major sucking pests on brinjal under field conditions. J. basic appl. sci., 13: 133-138. https://doi.org/10.6000/1927-5129.2017.13.22
- Alinejad M., Kheradmand K., Fathipour Y. 2016. Assessment of sublethal effects of spirodiclofen on biological performance of the predatory mite, Amblyseius swirskii. Syst. Appl. Acarol., 21: 375-384. https://doi.org/10.11158/saa.21.3.12
- Asadi P., Sedaratian-Jahromi A., Ghane-Jahromi M., Haghani M. 2019. How spiromesifen affects some biological parameters and switching behavior of predatory mite Amblyseius swirskii (acari: Phytoseiidae) when feeding on different ratios of mixed preys. Persian J. Acarol., 8: 239-251.
- Castro A.A., Lacerda M.C., Zanuncio T.V., Ramalho F.D.S., Polanczyk R.A., Serrao J.E., Zanuncio J.C. 2012. Effect of the insect growth regulator diflubenzuron on the predator Podisus nigrispinus (heteroptera: Pentatomidae). Ecotoxicology, 21: 96-103. https://doi.org/10.1007/s10646-011-0769-z
- Chi H. 1988. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ Entomol 17: 26-34. https://doi.org/10.1093/ee/17.1.26
- Chi H. 2020a. Consume-mschart: A computer program for predation rate study based on age-stage, two-sex life table http://140120197173/Ecology/
 .
. - Chi H. 2020b. Twosex-mschart: A computer program for the age-stage, two-sex life table analysis http://140120197173/Ecology/ .
- Chi H., Su H.Y. 2006. Age-stage, two-sex life tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and its host Myzus persicae (sulzer)(homoptera: Aphididae) with mathematical proof of the relationship between female fecundity and the net reproductive rate. Environ. Entomol., 35: 10-21. https://doi.org/10.1603/0046-225X-35.1.10
- Chi H., Yang T.C. 2003. Two-sex life table and predation rate of Propylaea japonica thunberg (Coleoptera: Coccinellidae) fed on Myzus persicae (Sulzer) (Homoptera: Aphididae). Environ. Entomol., 32: 327-333. https://doi.org/10.1603/0046-225X-32.2.327
- Chi H., You M., Atlihan R., Smith C.L., Kavousi A., Özgökçe M.S., Güncan A., Tuan S.J., Fu J.W., Xu Y.Y., Zheng F.Q. 2020. Age-stage, two-sex life table: An introduction to theory, data analysis, and application. Entomol. Gen., 40: 102-123. https://doi.org/10.1127/entomologia/2020/0936
- Cloyd R.A., Galle C.L., Keith S.R. 2006. Compatibility of three miticides with the predatory mites Neoseiulus californicus McGregor and Phytoseiulus persimilis Athias-Henriot (Acari: Phytoseiidae). Hort Science, 41: 707-710. https://doi.org/10.21273/HORTSCI.41.3.707
- Dai C., Ricupero M., Wang Z., Desneux N., Biondi A., Lu Y. 2021. Transgenerational effects of a neonicotinoid and a novel sulfoximine insecticide on the harlequin ladybird. Insects, 12: 681. https://doi.org/10.3390/insects12080681
- Dara S.K. 2017. Managing western flower thrips (Thysanoptera: Thripidae) on lettuce and green peach aphid and cabbage aphid (Hemiptera: Aphididae) on broccoli with chemical insecticides and the entomopathogenic fungus Beauveria bassiana (Hypocreales: Clavicipitaceae). Open Plant. Sci., J 10: 21-28. https://doi.org/10.2174/1874294701710010021
- De Castro A.A., Corrêa A.S., Legaspi J.C., Guedes R.N.C., Serrão J.E., Zanuncio, J.C. 2013. Survival and behavior of the insecticide-exposed predators Podisus nigrispinus and Supputius cincticeps (Heteroptera: Pentatomidae). Chemosphere, 93: 1043-1050. https://doi.org/10.1016/j.chemosphere.2013.05.075
- Dekeyser M.A. 2005. Acaricide mode of action. Pest Manag. Sci., 61: 103-110. https://doi.org/10.1002/ps.994
- Desneux, N., Decourtye A., Delpuech J.M. 2007. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol., 52: 81-106. https://doi.org/10.1146/annurev.ento.52.110405.091440
- Guedes R.N.C., Smagghe G., Stark J.D., Desneux, N. 2016. Pesticide-induced stress in arthropod pests for optimized integrated pest management programs. Annu. Rev. Entomol., 61: 43-62. https://doi.org/10.1146/annurev-ento-010715-023646
- Hamedi N., Fathipour Y., Saber M., Garjan A.S. 2009. Sublethal effects of two common acaricides on the consumption of Tetranychus urticae (Prostigmata: Tetranychidae) by Phytoseius plumifer (Mesostigmata: Phytoseiidae). Syst. Appl. Acarol., 14: 197-205. https://doi.org/10.11158/saa.14.3.4
- Kang Z.W., Liu F.H., Pang R.P., Tian H.G., Liu T.X. 2018. Effect of sublethal doses of imidacloprid on the biological performance of aphid endoparasitoid Aphidius gifuensis (Hymenoptera: Aphidiidae) and influence on its related gene expression. Front. Physiol., 9: 1729. https://doi.org/10.3389/fphys.2018.01729
- Kumari S., Chauhan U., Kumari A., Nadda G. 2017. Comparative toxicities of novel and conventional acaricides against different stages of Tetranychus urticae Koch (Acarina: Tetranychidae). J. Saudi Soc. Agric. Sci., 16: 191-196. https://doi.org/10.1016/j.jssas.2015.06.003
- Lee S.M., Kim S.S. 2015. Susceptibility of the predatory mite, Neoseiulus californicus (Acari: Phytoseiidae) to acaricides. Korean J. Pestic. Sci., 19: 418-423. https://doi.org/10.7585/kjps.2015.19.4.418
- Li Y.Y., Fan X., Zhang G.H., Liu Y.Q., Chen H.Q., Liu H., Wang J.J. 2017. Sublethal effects of bifenazate on life history and population parameters of Tetranychus urticae (Acari: Tetranychidae). Syst. Appl. Acarol., 22: 148-158. https://doi.org/10.11158/saa.22.1.15
- Lu Y., Wu K., Jiang Y., Guo Y., Desneux N. 2012. Widespread adoption of bt cotton and insecticide decrease promotes biocontrol services. Nature, 487: 362-365. https://doi.org/10.1038/nature11153
- Majidpour M., Maroofpour N., Ghane-Jahromi M., Guedes R.N.C. 2020. Thiacloprid+ deltamethrin on the life-table parameters of the cotton aphid, Aphis gossypii (Hemiptera: Aphididae), and the parasitoid, Aphidius flaviventris (Hymenoptera: Aphelinidae). J. Econ. Entomol., 113: 2723-2731. https://doi.org/10.1093/jee/toaa214
- Marcic D. 2007. Sublethal effects of spirodiclofen on life history and life-table parameters of two-spotted spider mite (Tetranychus urticae). Exp. Appl. Acarol., 42: 121-129. https://doi.org/10.1007/s10493-007-9082-1
- Marcic D., Ogurlic I., Mutavdzic S., Peric P. 2010. The effects of spiromesifen on life history traits and population growth of two-spotted spider mite (Acari: Tetranychidae). Exp. Appl. Acarol., 50: 255-267. https://doi.org/10.1007/s10493-009-9316-5
- Marcic D., Ogurlić I., Mutavdžić S., Perić P. 2009. The effect of spiromesifen on the reproductive potential of Tetranychus urticae Koch (Acari: Tetranychidae). Pestic. Fitomed, 24: 203-209. https://doi.org/10.2298/PIF0903203M
- Maroofpour N., Mousavi M., Hejazi M.J., Iranipour S., Hamishehkar H., Desneux N., Biondi A., Haddi K. 2021. Comparative selectivity of nano and commercial formulations of pirimicarb on a target pest, Brevicoryne brassicae, and its predator Chrysoperla carnea. Ecotoxicology, 30: 361-372. https://doi.org/10.1007/s10646-021-02349-x
- Maroufpoor M., Ghoosta Y., Pourmirza A.A., Lotfalizadeh H. 2016. The effects of selected acaricides on life table parameters of the predatory mite, Neoseiulus californicus fed on european red mite. North-West J. Zool., 12: 1-6. http://biozoojournals.ro/nwjz/index.html .
- McMurtry J.A., De Moraes G.J., Sourassou N.F. 2013. Revision of the life-styles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Syst. Appl. Acarol., 18: 297-320. https://doi.org/10.11158/saa.18.4.1
- Mollaloo M.G., Kheradmand K., Sadeghi R., Talebi A.A. 2017. Demographic analysis of sublethal effects of spiromesifen on Neoseiulus californicus (Acari: Phytoseiidae). Acarologia, 57: 571-580. https://doi.org/10.24349/acarologia/20174173
- Müller T., Gesing M.A., Segeler M., Müller C. 2019. Sublethal insecticide exposure of an herbivore alters the response of its predator. Environ. Pollut., 247: 39-45. https://doi.org/10.1016/j.envpol.2018.12.040
- Nauen,R., Schnorbach H. 2005. The biological profile of spiromesifen (Oberon)-a new tetronic acid insecticide/acaricide. Pflanzenschutz-Nachr Bayer, 58: 417-440.
- Phukan B., Rahman S., Bhuyan K.K. 2017. Effects of botanicals and acaricides on management of Tetranychus urticae (Koch) in tomato. J. Entomol. Zool. Stud., 5: 241-46. https://www.entomoljournal.com/archives/2017/vol5issue3/PartD/5-2-135-954.pdf .
- Rhodes E.M., Liburd O.E., Kelts C., Rondon S.I., Francis R.R. 2006. Comparison of single and combination treatments of Phytoseiulus persimilis, Neoseiulus californicus, and acramite (bifenazate) for control of twospotted spider mites in strawberries. Exp. Appl. Acarol., 39: 213-225. https://doi.org/10.1007/s10493-006-9005-6
- Sangak Sani N., Kheradmand K., Talebi, A.A. 2019. Sublethal effects of spirodiclofen on the demographic parameters of Tetranychus urticae Koch (Acari: Tetranychidae). Arch. Phytopathol. Plant Prot., 52: 938-952. https://doi.org/10.1080/03235408.2019.1668593
- Sarbaz S., Goldasteh S., Zamani A.A., Solymannejadiyan E., Vafaei Shoushtari R. 2017. Side effects of spiromesifen and spirodiclofen on life table parameters of the predatory mite, Neoseiulus californicus McGregor (Acari: Phytoseiidae). Int. J. Acarology, 43: 380-386. https://doi.org/10.1080/01647954.2017.1325396
- Sato M.E., Da Silva M.Z., Raga A., Cangani K.G., Veronez B., Nicastro R.L. 2011. Spiromesifen toxicity to the spider mite Tetranychus urticae and selectivity to the predator Neoseiulus californicus. Phytoparasitica, 39, 437-445. https://doi.org/10.1007/s12600-011-0189-x
- Shang S., Chang Y., Li W.Z., Chang-Qing W., Peng-Cheng N. 2022. Effects of b-azolemiteacrylic on life-history traits and demographic parameters of two-spotted spider mite, Tetranychus urticae (Acari: Tetranychidae). Exp. Appl. Acarol., 86: 61-71. https://doi.org/10.1007/s10493-021-00678-4
- Susurluk H., Gürkan M.O. 2020. Mode of inheritance and biochemical mechanisms underlying lambda-cyhalothrin and bifenthrin resistance in the laboratory-selected two-spotted spider mite, Tetranychus urticae. Crop Prot., 137: 105280. https://doi.org/10.1016/j.cropro.2020.105280
- Tuan S.J., Lee C.C., Chi H. 2014. Population and damage projection of Spodoptera litura (F.) on peanuts (Arachis hypogaea L.) under different conditions using the age-stage, two-sex life table. Pest Manag. Sci., 70: 805-813. https://doi.org/10.1002/ps.3618
- Wachendorff U., Nauen R., Schnorbach H.J., Stumpf N., Elbert A., 2002. The biological profile of spirodiclofen (envidor®) a new selective tetronic acid acaricide. Pflanzenschutz-Nachr Bayer, 55: 149-176.



2022-01-26
Date accepted:
2022-08-01
Date published:
2022-08-29
Edited by:
Tsolakis, Haralabos

This work is licensed under a Creative Commons Attribution 4.0 International License
2022 Rajaee, Fahimeh; Ghane-Jahromi, Mojtaba; Maroofpour, Nariman and Sedaratian-Jahromi, Amin
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)