




Climatic and cultivar effects on phytoseiid species establishment and seasonal abundance on citrus
Warburg, Sharon1 ; Gafni, Roni2 ; Inbar, Moshe3 ; Gal, Shira4 ; Palevsky, Eric5 and Sadeh, Asaf6
1Dept. of Entomology, Newe-Ya’ar Research Center, Agricultural Research Organization (ARO), P.O. Box 1021, Ramat Yishay 30095, Israel & Department of Evolutionary and Environmental Biology, University of Haifa, Haifa 31905, Israel.
2Dept. of Entomology, Newe-Ya’ar Research Center, Agricultural Research Organization (ARO), P.O. Box 1021, Ramat Yishay 30095, Israel & Department of Natural Resources, Institute of Plant Sciences, ARO, Volcani Center, P.O. Box 15159, Rishon Letsion 7505101, Israel.
3Department of Evolutionary and Environmental Biology, University of Haifa, Haifa 31905, Israel.
4Dept. of Entomology, Newe-Ya’ar Research Center, Agricultural Research Organization (ARO), P.O. Box 1021, Ramat Yishay 30095, Israel.
5✉ Dept. of Entomology, Newe-Ya’ar Research Center, Agricultural Research Organization (ARO), P.O. Box 1021, Ramat Yishay 30095, Israel.
6✉ Department of Natural Resources, Institute of Plant Sciences, ARO, Volcani Center, P.O. Box 15159, Rishon Letsion 7505101, Israel.
2019 - Volume: 59 Issue: 4 pages: 443-455
https://doi.org/10.24349/acarologia/20194346Original research
Keywords
Abstract
Introduction
Phytoseiids are mostly generalist predators that occur both in natural habitats and in crops (McMurtry et al., 2013). They can therefore potentially provide ecosystem services in agricultural pest control (Gerson et al., 2003). However, not all phytoseiid species are equally effective as biocontrol agents. In addition, negative and asymmetrical intraguild interactions such as intraguild predation (Rosenheim et al., 1995) and priority effects (Montserrat et al., 2008; van der Hammen et al., 2010) can limit the capacity of phytoseiid communities to suppress pests such as phytophagous mites (although the frequency of itraguild predation is debated; Guzmán et al., 2016). Furthermore, different environmental conditions, such as climate and host plant traits, may affect the relative dominance of different phytoseiid species in different habitats (McMurtry et al., 2013; Tixier, 2018), or alter key interspecific interactions (Urbaneja-Bernat et al., 2019) among them. Understanding the factors that influence mite community structure and dynamics across habitat types and seasons may help to manage ecosystem services and optimize biocontrol programs.
Overall forty six phytoseiid species have been collected from citrus in different Mediterranean countries (Vacante and Gerson, 2012). Euseius stipulatus Athias-Henriot was the most abundant species on citrus plants in most countries, except Israel, Egypt and Lebanon, where Amblyseius swirskii Athias-Henriot and Typhlodromus athiasae Porath and Swirski dominated (Rasmy et al., 1972; McMurtry, 1977; Vacante and Gerson, 2012; Abdelgayed et al., 2017). In Israel, phytoseiid species recorded in citrus orchards (Porath and Swirski, 1965) include Iphiseius degenerans Berlese and A. swirskii that are generally prevalent in orchards in the humid coastal region, T. athiasae that is more dominant in drier, inland regions, and Euseius scutalis Athias-Henriot, which is common on avocado (Maoz et al., 2011) but rare on citrus (Porath and Swirski, 1965).
A key cosmopolitan pest of citrus is the citrus rust mite (CRM) Phyllocoptruta oleivora Ashmead. A field study conducted by Smith and Papacek (1991) in Australia demonstrated that better control of P. oleivora and of the brown citrus rust mite Tegolophus australis Keifer can be achieved with Euseius victoriensis, using a windborne pollen-releasing Rhodes grass Chloris gayana Kunth cover crop. With the aim of establishing new biocontrol agents for CRM control, five phytoseiid species, among them E. stipulatus were imported and released during 1994-1996 in all citrus growing regions of Israel (Argov et al., 2002). Studies conducted in the laboratory with citrus seedlings provisioned with pollen as supplemental food, indicated that the indigenous E. scutalis could be an effective CRM biocontrol agent. In contrast, A. swirskii, an intraguild predator of E. scutalis, had no effect on CRM control (Maoz et al., 2016). Thus, the success of biological control may depend critically on the phytoseiid community structure and seasonal dynamics, particularly of these two functionally distinct genera.
Warburg et al. (2019) followed the dynamics and abundance relationships between released E. stipulatus and other naturally occurring phytoseiids at varying distances from Rhodes grass cover crop strips in two orchards in Israel's coastal plain; a Shamouti Citrus sinensis (L.) Osbeck orchard, and a pomelo Citrus grandis (L.) Osbeck orchard. In the pomelo orchard, E. stipulatus did not establish, and A. swirskii was found to be the dominant species from May through October. In contrast, in the Shamouti orchard, E. stipulatus was the dominant species in April and May. During June and July, the densities of all phytoseiids were extremely low, but E. stipulatus re-established from August through October, co-existing with A. swirskii at similar densities. In both orchards, pollen provisioning next to the Rhodes grass strips resulted in substantially higher phytoseiid population densities. However, negative relationships between E. stipulatus and A. swirskii (possibly due to intraguild predation) were observed only in the Pomelo orchard. In the Shamouti orchard, no such relationship was detected, E. stipulatus densities were high, and a significant effect on CRM control was observed.
We hypothesized that seasonal phytoseiid community structure in orchards is affected by cultivar. We also hypothesized that temperature and cultivar affect the relative performance of different mite species, specifically E. stipulatus and A. swirskii. To test these hypotheses, we evaluated phytoseiid establishment on potted seedlings of several cultivars, from spring through mid-summer, with and without pollen provisioning. To determine the potential role of fine-scale cultivar effects that might be mediated by leaf morphological or chemical traits, we also conducted controlled laboratory experiments on leaf discs of the two previously studied cultivars, pomelo and Shamouti (Warburg et al., 2019). These experiments evaluated the survival and reproduction of A. swirskii and E. stipulatus on each cultivar, under daily-fluctuating temperature regimes that simulated the spring and mid-summer seasons in the coastal plain of Israel.
Materials and methods
General methods
Experiments were performed at the Newe-Ya'ar Research Center (32°70'N; 35°18'E), located in the Jezreel Valley. A pollen mix from Mount Tabor oak Quercus ithaburensis, cattail Typha domingensis and maize Zea mays (in even parts) was used as a source of supplementary food in all experiments. Oak pollen was collected from trees in Newe-Ya'ar in March, cattail pollen from plants growing on drainage ditch banks near Newe-Ya'ar in May, and maize pollen was collected from commercial maize fields in June, before any insecticide sprays were applied. The flowers bearing pollen grains were dried for a week on paper. Dried flowers and loose pollen grains were sieved (180 μm sieve) and stored at -20 °C. Mites that were used in both experiments (E scutalis and E. stipulatus in the seedling experiment, and A. swirskii and E. stipulatus in the lead disc experiment) were received from Bio-Bee, Sdeh Eliyahu, Israel (http://www.biobee.com ![]() ).
).
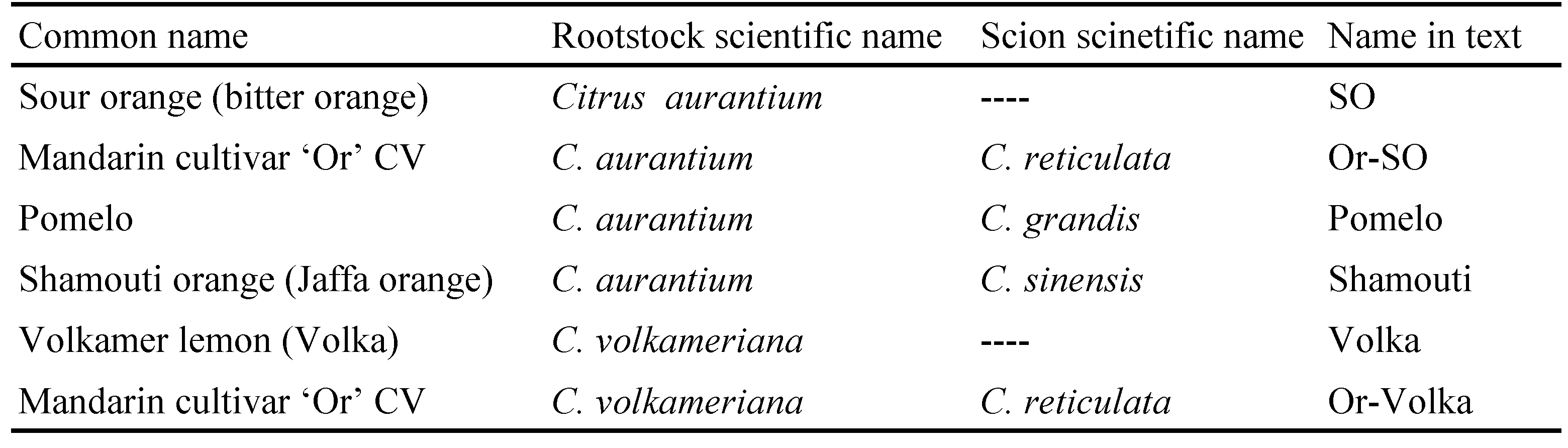
Outdoor seedling experiment
Eighteen seedlings (1 year old trees) of the following six rootstock / rootstock-scion combinations (hereafter referred to simply as ''cultivars''), were purchased from the Messilot nursery (https://www.messilot.net/greenhouse ![]() ): Sour orange (SO), Or grafted on SO (Or-SO), Pomelo grafted on SO (Pomelo), Shamouti grafted on SO (Shamouti), Volka, Or grafted on Volka (Or-Volka) (for full common and Latin names of citrus rootstock and scions see Table 1). Seedlings were approximately 1.5 m high when transplanted to ten liter pails and arranged randomly on a levelled gravel surface at least one meter apart. The seedlings were irrigated and fertilized following standard practice.
): Sour orange (SO), Or grafted on SO (Or-SO), Pomelo grafted on SO (Pomelo), Shamouti grafted on SO (Shamouti), Volka, Or grafted on Volka (Or-Volka) (for full common and Latin names of citrus rootstock and scions see Table 1). Seedlings were approximately 1.5 m high when transplanted to ten liter pails and arranged randomly on a levelled gravel surface at least one meter apart. The seedlings were irrigated and fertilized following standard practice.

Seedlings were randomly assigned to three predator and pollen treatments: E. scutalis release + pollen augmentation, E. stipulatus release + pollen augmentation, and control (no predators or pollen added), with 6 replicates per treatment. Rearing of E. scutalis and E. stipulatus was done on paper arenas (Argov et al., 2002) with maize pollen provided by Bio-Bee (http://www.biobee.com ![]() ). Predator release was performed once, on March 17, 2016, by fastening a 1 ml pipette tip containing 200 motile stages (or empty tips in the control) to each seedling. To facilitate the establishment of released predators, a pollen mix (described in the general methods) was added with a fine forceps on seedling leaves, starting at the time of predator release, and subsequently three times per week throughout the experiment. Seedlings were sampled for phytoseiid abundances in April, in June, and in August, by beating the top, middle and bottom of the seedling (three times each) with a section of flexible plastic irrigation pipe, being careful not to damage the seedling. Mites were collected from the beating tray with aspirators prepared from 200 µl micropipette tips (Argov et al., 2002). Collected mites were cleared, mounted in Hoyer medium and identified to the species level (using the description provided in Papadoulis et al., 2009). The spring sampling, in April, was indicative of cultivar compatibility with the establishment of the released predatory species, and with the colonization of other phytoseiid species from the surroundings natural environment, with and without pollen provisioning. The early summer sampling, in June, allowed to evaluate the persistence of the established phytoseiid species on the different cultivars during a season characterized by extreme events of high temperature and low humidity that resulted in phytoseiid population collapses in previous field studies (Warburg et al., 2019). Finally, the mid-summer sampling, in August, several weeks after humidity levels typically increase and stabilize, tested the compatibility of cultivars to re-establishment by phytoseiid species from the surrounding natural environment, with and without pollen provisioning.
). Predator release was performed once, on March 17, 2016, by fastening a 1 ml pipette tip containing 200 motile stages (or empty tips in the control) to each seedling. To facilitate the establishment of released predators, a pollen mix (described in the general methods) was added with a fine forceps on seedling leaves, starting at the time of predator release, and subsequently three times per week throughout the experiment. Seedlings were sampled for phytoseiid abundances in April, in June, and in August, by beating the top, middle and bottom of the seedling (three times each) with a section of flexible plastic irrigation pipe, being careful not to damage the seedling. Mites were collected from the beating tray with aspirators prepared from 200 µl micropipette tips (Argov et al., 2002). Collected mites were cleared, mounted in Hoyer medium and identified to the species level (using the description provided in Papadoulis et al., 2009). The spring sampling, in April, was indicative of cultivar compatibility with the establishment of the released predatory species, and with the colonization of other phytoseiid species from the surroundings natural environment, with and without pollen provisioning. The early summer sampling, in June, allowed to evaluate the persistence of the established phytoseiid species on the different cultivars during a season characterized by extreme events of high temperature and low humidity that resulted in phytoseiid population collapses in previous field studies (Warburg et al., 2019). Finally, the mid-summer sampling, in August, several weeks after humidity levels typically increase and stabilize, tested the compatibility of cultivars to re-establishment by phytoseiid species from the surrounding natural environment, with and without pollen provisioning.
Leaf disc experiment
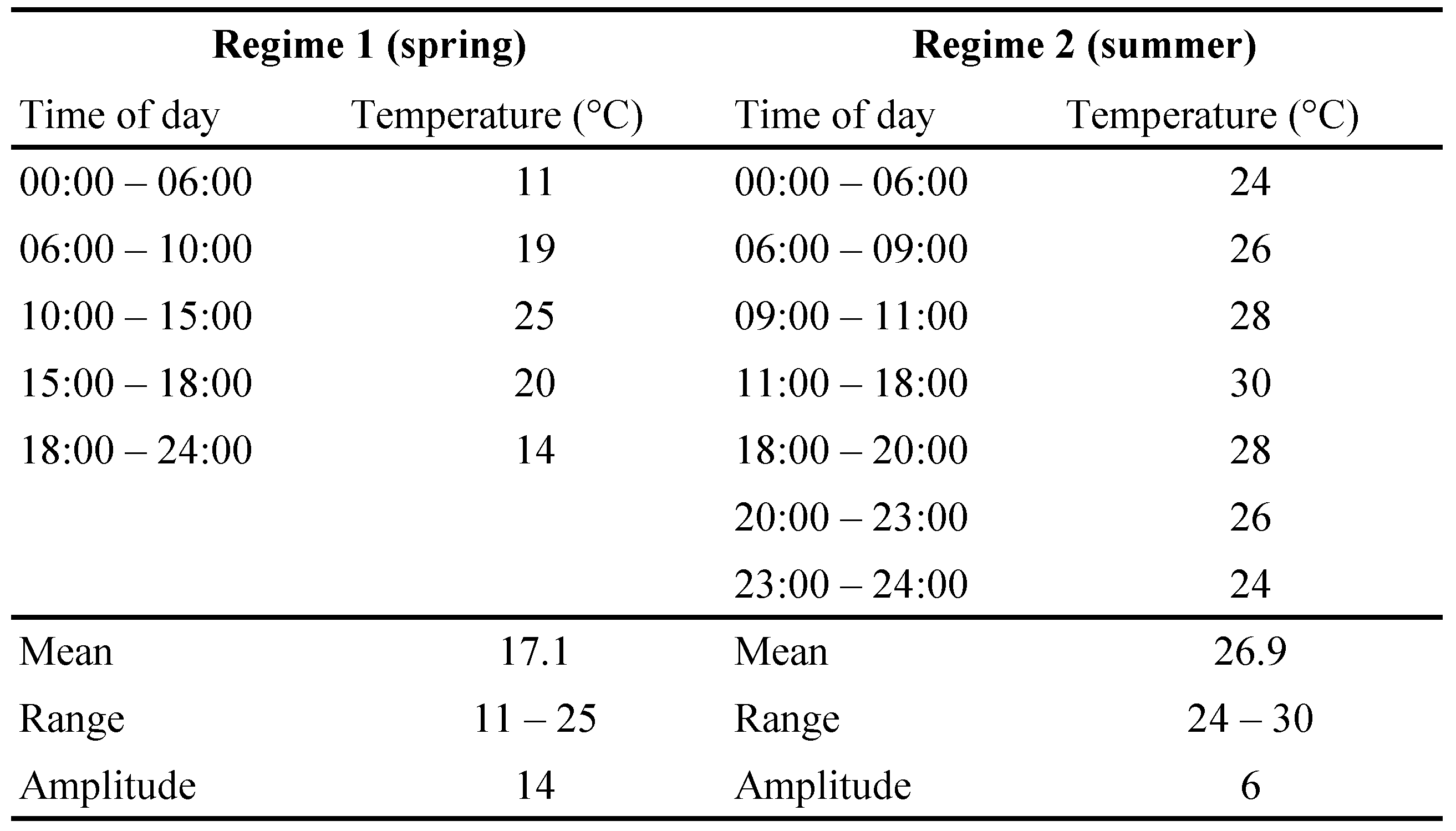
To evaluate potential cultivar leaf effects on the survival and reproductive success of A. swirskii and E. stipulatus under different climatic conditions, we conducted an experiment in climate-controlled growth chambers (https://www.conviron.com/products/a1000-reach-in-plant-growth-chamber ![]() ). The chambers were programed to follow two regimes based on typical daily temperature fluctuations in the spring (April) and mid-summer (July-August) in the coastal plain of Israel (Table 2). Rearing of E. stipulatus and A. swirskii was done at 24 ± 5 °C at Newe Ya'ar, using the pollen mix described above. Prior to each experiment, polyester fibers as substrate for egg laying were placed on rearing arenas of E. stipulatus and A. swirskii for 24 hr. Fibers with eggs were then placed on similar rearing units in each growth chamber and allowed to develop, mate and oviposit, thus rearing a full generation under the experimental temperature regimes. Second-generation eggs of each species at standardized ages (0–1 day old) were then collected and placed in groups of ten on leaf discs (20 mm in diameter) of either pomelo or Shamouti. Each leaf disc was floated on polyacrylamide gel in a 120 ml container, and replicated 6–10 times for each species by cultivar combination, under each temperature regime. Pollen mix was applied on each leaf disc twice per week with a fine forceps. On the day following the onset of oviposition in each temperature regime (days 6 and 13 in the summer and spring regimes, respectively), we recorded the number of females in each of the containers. Eggs oviposited during the experiment were counted and removed daily until the experiment was terminated (on days 10 and 21, in the summer and spring regimes, respectively). We calculated the mean daily reproductive output per female for each container as the total number of eggs oviposited in the container divided by the number of females observed at the onset of oviposition and by the number of reproductive days (5 days in the summer regime, and 9 days in the spring regime). This measure of mite fitness, or net contribution to population growth, incorporates variation in multiple underlying factors that were not recorded in the experiment, such as in female fecundity and survival, sex ratio, and mating success. In addition, mean survival per container was calculated for all individuals (regardless of sex) through the entire duration of the experiment.
). The chambers were programed to follow two regimes based on typical daily temperature fluctuations in the spring (April) and mid-summer (July-August) in the coastal plain of Israel (Table 2). Rearing of E. stipulatus and A. swirskii was done at 24 ± 5 °C at Newe Ya'ar, using the pollen mix described above. Prior to each experiment, polyester fibers as substrate for egg laying were placed on rearing arenas of E. stipulatus and A. swirskii for 24 hr. Fibers with eggs were then placed on similar rearing units in each growth chamber and allowed to develop, mate and oviposit, thus rearing a full generation under the experimental temperature regimes. Second-generation eggs of each species at standardized ages (0–1 day old) were then collected and placed in groups of ten on leaf discs (20 mm in diameter) of either pomelo or Shamouti. Each leaf disc was floated on polyacrylamide gel in a 120 ml container, and replicated 6–10 times for each species by cultivar combination, under each temperature regime. Pollen mix was applied on each leaf disc twice per week with a fine forceps. On the day following the onset of oviposition in each temperature regime (days 6 and 13 in the summer and spring regimes, respectively), we recorded the number of females in each of the containers. Eggs oviposited during the experiment were counted and removed daily until the experiment was terminated (on days 10 and 21, in the summer and spring regimes, respectively). We calculated the mean daily reproductive output per female for each container as the total number of eggs oviposited in the container divided by the number of females observed at the onset of oviposition and by the number of reproductive days (5 days in the summer regime, and 9 days in the spring regime). This measure of mite fitness, or net contribution to population growth, incorporates variation in multiple underlying factors that were not recorded in the experiment, such as in female fecundity and survival, sex ratio, and mating success. In addition, mean survival per container was calculated for all individuals (regardless of sex) through the entire duration of the experiment.
Statistical analyses
Statistical analyses were done in R 3.5.1 statistical computing software (R Core Team, 2018). To analyze phytoseiid abundances in April in the seedlings experiment, we used general linear models (LM) for E. stipulatus and I. degenerans, and generalized linear models (GLM) with negative binomial error distributions and a log link function for A. swirskii and T. athiasae, with predator release treatment and cultivar as predictors. In August, we used LM for A. swirskii abundances, and GLM (negative binomial, log link) for T. athiasae abundances. In the August analyses, we considered our treatments to contrast the presence vs. absence of pollen augmentation, because the released E. stipulatus and E. scutalis populations had disappeared in June without recovering. In modeling T. athiasae abundance in August, we included the abundances of its intraguild predator, A. swirskii (Maoz et al., 2016), as a predictor, and removed the pollen × cultivar interaction term to maintain statistical power. In cases of heteroscedasticity of model residuals, we used the HC3 heteroscedastic-consistent covariance matrix method (Zeileis, 2004) in ANOVAs and post-hoc comparisons.
In the leaf discs experiment, female reproduction was modeled as a general linear model of species and cultivar under each temperature regime. Mite survival throughout the experiment was modeled as a GLM of species and cultivar under each temperature regime, with binomially-distributed errors and logit link function.
Results
Outdoor seedling experiment
The results of the statistical analyses used for this experiment are presented in Table 3.

In April — five weeks post release, E. stipulatus was recovered in substantial abundances (mean ± SE: 15.0 ± 2.1 individuals per sample) on seedlings on which it was released, whereas very few mites of this species were found on the other seedlings (mean ± SE: 1.85 ± 0.34; p < 0.0001). The cultivars had no significant effect (p = 0.671), and no significant interaction between cultivar and predator release was observed (p = 0.821) (Figure 1A). In contrast, despite its recent release, abundances of E. scutalis were very low on all seedlings (mean: 0.37, range: 0–6 individuals per sample), and it was found almost exclusively on seedlings on which it was released.
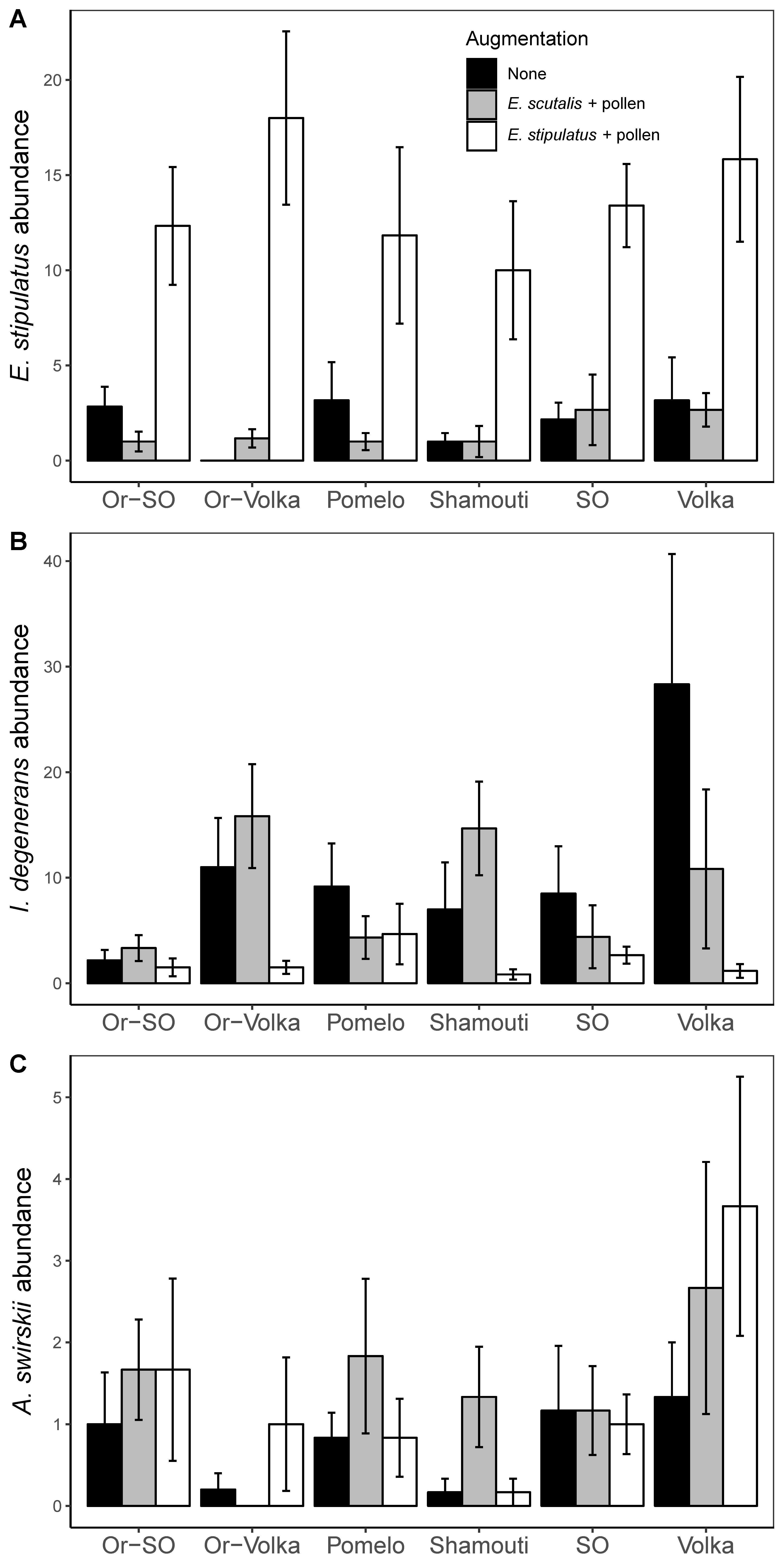
Colonizing I. degenerans established in high abundances on seedlings without release (mean ± SE: 11.03 ± 2.77 individuals per sample), and on E. scutalis-augmented seedlings (mean ± SE: 11.97 ± 3.47 individuals per sample). It generally had lower abundances on seedlings where E. stipulatus was released (mean ± SE: 2.06 ± 0.55 individuals per sample), despite the provisioning of pollen (p = 0.010; Tukey's comparisons between E. stipulatus augmentation and no augmentation treatments t = 3.152; p = 0.006; between E. stipulatus augmentation and E. scutalis augmentation treatments t = 3.337; p = 0.003) (Figure 1B). Whereas the main effect of cultivar on I. degenerans abundance was non-significant (p = 0.362), the negative effect of E. stipulatus on the establishment of I. degenerans appeared mostly on Shamouti-SO, Volka and Or-Volka and was not consistent across all cultivars (p = 0.054).
Relatively few colonizing A. swirskii were recovered on all seedlings (mean: 1.21, range: 0 – 11 individuals per sample), without significant effects of predator augmentation (p = 0.141), but with a significant cultivar effect (p = 0.001), with lower densities on Or-Volka and on Shamouti than on the other cultivars (Figure 1C).
Abundances of T. athiasae were also low on all seedlings (mean: 1.71, range: 0 – 9 individuals per sample), and not affected by phytoseiid augmentation or by cultivar (Table 3).
In June — only two specimens of E. stipulatus, two specimens of E. scutalis, and no I. degenerans were recovered from all the seedlings. Abundances of A. swirskii (mean: 0.43, range: 0 – 7 individuals per sample) and of T. athiasae (mean: 1.73, range: 0–6 individuals per sample) were reduced compared to April, or remained similarly low, without any cultivar or augmentation treatment effects.
In August — the abundances of E. stipulatus and E. scutalis remained negligible (mean: 0.17 and 0.27, range: 0–5 and 0–10 individuals per sample, respectively). Similar to June, no I. degenerans mites were detected in August. In contrast, abundances of A. swirskii were very high, especially on the seedlings that were provisioned with pollen (compared to unprovisioned seedlings, p = 0.004) (Figure 2A). There was also a significant cultivar effect (p=0.006), with weakest A. swirskii establishment on pomelo and Shamouti, and strongest establishment on both rootstocks (Sour and Volka) and on Or-Volka.

The abundance of T. athiasae also increased substantially from June to August, but still remained low compared to A. swirskii abundance. There was a significant negative relationship between T. athiasae abundances and those of A. swirskii (p<0.0001), its intraguild predator (Maoz et al., 2016). In addition, cultivar has a significant effect (p = 0.031), with the greatest difference occurring between Pomelo and Shamouti (Figure 2B). Pollen provisioning did not have a significant impact on T. athiasae abundance (p = 0.123).
Throughout the experiment, no citrus arthropod pests were observed, probably due to the absence of nearby orchards.
Leaf disc experiment
Female reproductive output, combining female survival and fecundity, was much lower in the temperature regime simulating spring conditions (Figure 3A) than in the summer regime (Figure 3B), for both phytoseiid species and cultivars, despite the shorter experimental period of the latter regime. Under the spring regime, E. stipulatus reproductive output outperformed that of A. swirskii three-fold on Pomelo and six-fold on Shamouti (LM: species effect F1,31 = 19.436; p = 0.0001), without significant difference between cultivars (cultivar effect F1,31 = 0.007; p = 0.932). In contrast, under the summer regime, A. swirskii significantly outperformed E. stipulatus on both cultivars (LM: species effect F1,29 = 10.460; p = 0.003), without a significant difference between cultivars (cultivar effect F1,29= 0.249; p=0.621).
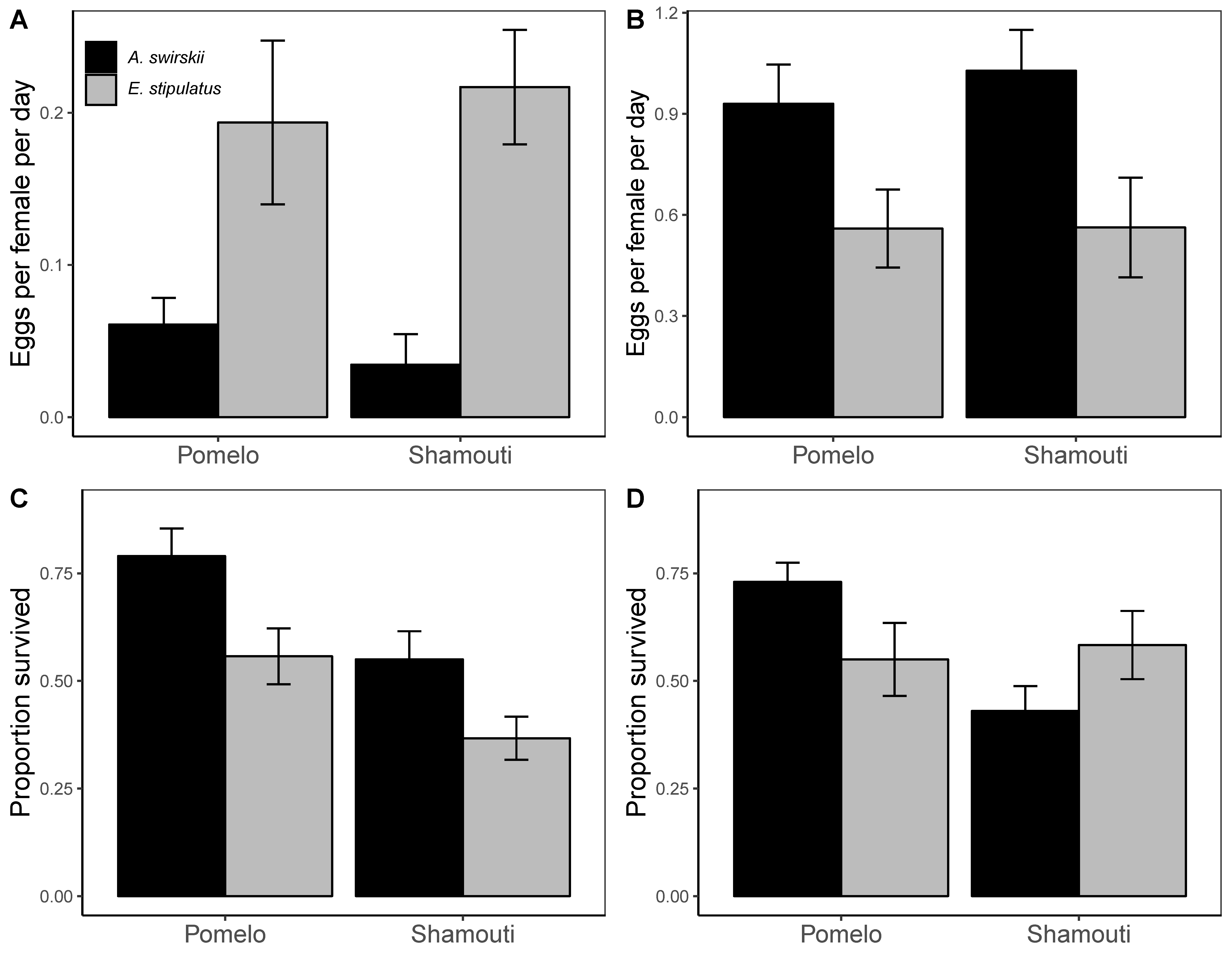
Survival of A. swirskii mites (of both sexes) under the spring regime was greater than that of E. stipulatus on both cultivars (Figure 3C; GLM: species effect χ2 = 16.304; p<0.0001), and both species showed greater survival on Pomelo than on Shamouti (cultivar effect χ2 = 18.456; p < 0.0001). Under the summer regime, A. swirskii survival was greater than that of E. stipulatus on Pomelo, but lower than that of E. stipulatus on Shamouti (Figure 3D; GLM: species×cultivar interaction χ2 = 8.853; p = 0.003).
Discussion
This study combined an outdoor seedlings experiment and a laboratory leaf disc experiment, both focusing on the roles of citrus cultivars and climatic conditions on the seasonal performance and abundances of introduced and naturally occurring phytoseiid species. The continuous pollen augmentation to some of the seedlings resembled orchards where pollen is either augmented manually (Biobest Belgium, 2013), or via wind-pollinated cover crops (Warburg et al., 2019), to support phytoseiid populations.
Phytoseiid communities on citrus during spring
The outdoor seedling experiment showed that augmented E. stipulatus population established and persisted through spring, regardless of cultivar (Figure 1A). However, the lack of E. stipulatus dispersal from where they were released to other seedlings seems to indicate limited dispersal of the augmented mites. Provisioning of pollen did not seem to be a limiting factor because of the large amounts of pollen shed from the nearby oak trees during late March and early April. Indeed, I. degenerans, a naturally-occurring species classified as a pollen feeding generalist (McMurtry et al., 2013), was found in high abundance on seedlings without pollen supplementation. Euseius stipulatus appeared to locally reduce I. degenerans colonization on some of the cultivars, likely due to asymmetrical intraguild interactions or priority effects between these species (Montserrat et al., 2008; van der Hammen et al., 2010). This appeared to be more prominent on some citrus cultivars than on others (namely, on Shamouti, Volka, and Or-Volka, but not on Sour orange, Or-SO, and Pomelo). This intraguild interaction coupled with the cultivar effect might be higher in commercial orchards where cultivar monocultures preclude spatial heterogeneity and small-scale source-sink dynamics among I. degenerans populations. Indeed, I. degenerans occurred at very low abundances in the Shamouti orchard studied by Warburg et al. (2019), and was far less prevalent than E. stipulatus in the spring. These results differ from those of a recent laboratory study (Calabuig et al., 2018), where gravid females of I. degenerans, with and without pollen, consumed substantially more heterospecific larvae than gravid females of E. stipulatus. Maoz et al. (2016) also reported very strong predation by I. degenerans on heterospecific (both E. scutalis and A. swirskii) larvae in leaf disc arena experiments. Thus, it seems that in outdoor and orchard conditions, or as a result of priority effects, the laboratory-observed direction of intraguild predation is either reversed or outweighed by a counteracting factor, leading to the displacement of I. degenerans by E. stipulatus.
Colonization by A. swirskii in the spring sampling (April) occurred in extremely low densities (Figure 1C), mirroring the springtime (March-May) low abundances of this predator in the large scale field experiments in both Pomelo and Shamouti orchards conducted by Warburg et al (2019). Under the temperature regime simulating spring conditions, the reproductive output of E. stipulatus females was much greater than that of A. swirskii females (Figure 3A), despite greater survival of A. swirskii (Figure 3C). Other laboratory studies conducted at a constant temperature of 18 °C reported similar differences between these two species both in productivity and in fecundity (Ferragut et al., 1987; Lee and Gillespie, 2011). However, survival in the leaf disc experiment reported herein may have been high mainly among the males, whose abundance contributes little to female reproductive success and to population dynamics.
Collapse of phytoseiid communities during early summer
The collapse of populations of all phytoseiid species in June also mirrors the dynamics observed by Warburg et al. (2019) in the commercial orchards. The population collapse in the current seedlings experiment likely occurred due to low humidity, in combination with high temperatures; the driest conditions were recorded in May, with a total of forty hours below 30% relative humidity during the fortnight preceding the June sampling, whereas relative humidity never dropped below 30% during the fortnight preceding the August sampling. Reports in the literature show that E. stipulatus is sensitive to both temperature and relative humidity (Ragusa, 1986; Ferragut et al., 1987; Sahraoui et al., 2014); In Sicily, E. stipulatus populations collapsed after four hours of temperatures above 40 °C, coupled with strong dry winds (Ragusa, 1986). Similar reductions in E. stipulatus populations occurred in Tunisia during July-August, when temperatures exceed 30–35 °C (Sahraoui et al., 2014), and in Spanish citrus orchards, after several days with temperature exceeding 34–38 °C and 30–40% RH (Ferragut et al., 1987). Thus, in the absence of viable populations of E. stipulatus in the surrounding habitat that can serve as sources of orchard re-colonization, early summer weather in Israel can be an obstacle for effective CRM biocontrol with this phytoseiid species.
Phytoseiid communities on citrus during mid-summer
By August, none of the species that were abundant during spring had rebounded, and they have been succeeded by naturally occurring populations of A. swirskii and T. athiasae (Figure 2). The nearly complete absence of E. stipulatus differs from the densities observed in the commercial (primarily Shamouti) orchards (Warburg et al., 2019), and may result from three, non-mutually exclusive factors: (i) reduced availability of refugia from the extreme climatic conditions in early summer on the small seedlings, compared to fully grown trees, (ii) suppression by the dominant intraguild predator, A. swirskii, as was also indicated by the patterns observed in the commercial Pomelo orchard (but not in the Shamouti orchard) (Warburg et al., 2019), and (iii) the higher reproductive output of A. swirskii under summer temperatures (Figure 3B).
A greater abundance of A. swirskii was found in August on seedlings where pollen was augmented than those that were not augmented, and the abundances of this species were significantly different among cultivars. A negative relationship was evident between T. athiasae and A. swirskii abundances (Figure 2B), as observed also in the commercial Pomelo orchard (Warburg et al., 2019), likely due to intraguild predation (Maoz et al., 2016).
The success of Euseius spp. introductions
After its introduction in 1994 (Argov et al., 2002), E. stipulatus was recovered for the first time in Northern Israel adjacent to its release site in 2011 (Maoz et al., 2016), and then in the coastal region approximately 10 km from their closest release sites in 2013 and 2014 (Warburg, unpublished). As far as we know, Warburg et al. (2019) was the first to report coexistence of A. swirskii and E. stipulatus on citrus in the field in Israel. In Egypt and Saudi Arabia (Elmoghazy, 2016; Abdelgayed et al., 2017) A. swirskii is abundant and E. stipulatus has not been reported. In the other citrus growing countries in the Mediterranean, E. stipulatus is the prevalent species (Abad-Moyano et al., 2009; Faraji et al., 2011; Tsagkarakis et al., 2011; Vacante and Gerson, 2012; Barbar, 2013; Satar et al., 2013) without record of A. swirskii. We believe that it is important to continue to follow the regional establishment of this important introduced phytoseiid species.
Interestingly, in the current seedlings experiment, E. scutalis found in abundance on citrus in other Mediterranean countries (Bounfour and McMurtry, 1987; Barbar, 2013; Barbar, 2014), failed to establish on any of the cultivars, despite releases in large numbers, pollen provisioning, and the lack of dominant intraguild predators in the spring, at the time of augmentation. A similar observation was made in our previous field experiment in Shamouti (Warburg et al., 2019), where, remarkably, high abundances of E. scutalis were found on adjacent avocado trees. Thus, while E. scutalis proved to be easily enhanced by Rhodes grass pollen and efficient in controlling spider mites on avocado (Maoz et al., 2011), it repeatedly proves to be unsuccessful on citrus in Israel.
Conclusions, prospects, and implications
Successful establishment of introduced phytoseiid species for biocontrol may critically depend on taking into account seasonal and climatic conditions in the orchard. Specifically, E. stipulatus is sensitive to hot and dry conditions that typically occur in early summer, in Israel, resulting in severe declines or extinction of augmented populations. This can potentially be mitigated proactively, by providing humid refugia such as cover crop strips or perennial plants in orchard margins. Introductory mite releases can be considered earlier in the spring, when E. stipulatus performs well, to allow the establishment of more robust populations prior to the early-summer period of adverse conditions.
The performance of E. stipulatus did not appear to be directly affected by citrus cultivars. However, our results suggest that the naturally-occurring phytoseiids may be more sensitive to cultivar effects, potentially also affecting E. stipulatus indirectly. Specifically, A. swirskii performance differed between cultivars in our seedling experiment, although not on the leaf discs, suggesting the relevant scale of cultivar-specific factors may be the tree level. In the current experiment, E. stipulatus did not co-occur with A. swirskii, and the laboratory experiment was not designed to test for interspecific interactions among the phytoseiids. However, a key observation in Warburg et al. (2019) was the indication of negative intraguild interactions between A. swirskii and E. stipulatus only in the Pomelo orchard, but not in the Shamouti orchard. Moreover, in the current outdoor experiment, cultivar-dependent negative interactions appeared to occur between E. stipulatus and I. degenerans, and between A. swirskii and T. athiasae. These may have implications for the control of different pest species that these phytoseiids can suppress. Further research should explicitly examine cultivar and temperature effects on the behavioral responses of phytoseiid species pairs, investigating, for instance, the expression of intraguild predation and oviposition habitat selection.
Acknowledgements
We thank the R&D staff at BioBee (http://www.biobee.com ![]() ) for their technical support and for providing the predatory mites for the releases. This work was supported by the Chief Scientist of the Israeli Ministry of Agriculture, research project 20-01-0102, awarded to AS and EP. This manuscript is a contribution of the Institute of Plant Protection and the Institute of Plant Sciences, Volcani Center, ARO, Israel.
) for their technical support and for providing the predatory mites for the releases. This work was supported by the Chief Scientist of the Israeli Ministry of Agriculture, research project 20-01-0102, awarded to AS and EP. This manuscript is a contribution of the Institute of Plant Protection and the Institute of Plant Sciences, Volcani Center, ARO, Israel.
References
Abad-Moyano R., Pina T., Dembilio O., Ferragut F., Urbaneja A. 2009. Survey of natural enemies of spider mites (Acari: Tetranychidae) in citrus orchards in eastern Spain. Experimental and Applied Acarology, 47: 49-61. doi:10.1007/s10493-008-9193-3 ![]()
Abdelgayed A., Negm M., Eraky S., Helal T., Moussa F. 2017. Phytophagous and predatory mites inhabiting citrus trees in Assiut Governorate, Upper Egypt. Assiut Journal of Agricultural Sciences, 48: 173-181. doi:10.21608/ajas.2016.3739 ![]()
Argov Y., Amitai S., Beattie G.A.C., Gerson U. 2002. Rearing, release and establishment of imported predatory mites to control citrus rust mite in Israel. BioControl, 47: 399-409. doi:10.1023/A:1015634813723 ![]()
Barbar Z. 2013. Survey of phytoseiid mite species (Acari: Phytoseiidae) in citrus orchards in Lattakia governorate, Syria. Acarologia, 53: 247-261. doi:10.1051/acarologia/20132098 ![]()
Barbar Z. 2014. Occurrence, population dynamics and winter phenology of spider mites and their phytoseiid predators in a citrus orchard in Syria. Acarologia, 54: 409-423. doi:10.1051/acarologia/20142143 ![]()
Biobest Belgium N.V. Biobest introduces Dyna-Mite®: a new predatory mite strategy in rose [Internet]. Westerlo: Biobest Belgium N.V.; [cited]. Available from: http://www.biobest.be/nieuws/289/3/0/ ![]() .
.
Bounfour M., McMurtry J.A. 1987. Biology and ecology of Euseius scutalis (Athias-Henriot) (Acarina: Phytoseiidae). Hilgardia, 55: 1-23. doi:10.3733/hilg.v55n05p023 ![]()
Calabuig A., Pekas A., Wäckers F. 2018. The quality of nonprey food affects cannibalism, intraguild predation, and hyperpredation in two species of phytoseiid mites. Journal of Economic Entomology 111: 72-77. doi:10.1093/jee/tox301 ![]()
Elmoghazy M. 2016. Survey and taxonomy of mites associated with fruit orchards trees from Sakaka governorate, Kingdom of Saudi Arabia. International Journal of Agriculture and Biosciences, 5: 341-346.
Faraji F., Çobanoğlu S., Çakmak I. 2011. A checklist and a key for the Phytoseiidae species of Turkey with two new species records (Acari: Mesostigmata). International Journal of Acarology, 37: 221-243. doi:10.1080/01647954.2011.558851 ![]()
Ferragut F., Garcia-Mari F., Costa-Comelles J., Laborda R. 1987. Influence of food and temperature on development and oviposition of Euseius stipulatus and Typhlodromus phialatus (Acari: Phytoseiidae). Experimental and Applied Acarology, 3: 317-329. doi:10.1007/BF01193168 ![]()
Gerson U., Smiley R.L., Ochoa R. 2003. Mites (Acari) for pest control. Oxford, UK: Blackwell Science. pp. 560. doi:10.1002/9780470750995 ![]()
Guzmán C., Sahún R., Montserrat M. 2016. Intraguild predation between phytoseiid mite species might not be so common. Experimental and Applied Acarology, 68: 441-453. doi:10.1007/s10493-015-9997-x ![]()
Lee H.S., Gillespie D.R. 2011. Life tables and development of Amblyseius swirskii (Acari: Phytoseiidae) at different temperatures. Experimental and applied acarology, 53: 17-27. doi:10.1007/s10493-010-9385-5 ![]()
Maoz Y., Gal S., Argov Y., Coll M., Palevsky E. 2011. Biocontrol of persea mite, Oligonychus perseae, with an exotic spider mite predator and an indigenous pollen feeder. Biological Control, 59: 147-157. doi:10.1016/j.biocontrol.2011.07.014 ![]()
Maoz Y., Gal S., Argov Y., Domeratzky S., Coll M., Palevsky E. 2016. Intraguild interactions among specialized pollen feeders and generalist phytoseiids and their effect on citrus rust mite suppression. Pest Management Science, 72: 940-949. doi:10.1002/ps.4073 ![]()
McMurtry J.A. 1977. Some predaceous mites (Phytoseiidae) on citrus in the Mediterranean region. Entomophaga, 22: 19-30. doi:10.1007/BF02372986 ![]()
McMurtry J.A., de Moraes G.J., Sourassou N.F. 2013. Revision of the lifestyles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Systematic and Applied Acarology, 18: 297-320. doi:10.11158/saa.18.4.1 ![]()
Montserrat M., Magalhães S., Sabelis M.W., de Roos A.M., Janssen A. 2008. Patterns of exclusion in an intraguild predator-prey system depend on initial conditions. Journal of Animal Ecology, 77: 624-630. doi:10.1111/j.1365-2656.2008.01363.x ![]()
Papadoulis G.T., Emmanouel N.G., Kapaxidi E.V. 2009. Phytoseiidae of Greece and Cyprus (Acari: Mesostigmata). Michigan, USA: Indira Publishing House. pp. 200.
Porath A., Swirski E. 1965. A survey of phytoseiid mites (Acarina: Phytoseiidae) on citrus, with a description of one new species. Israel Journal of Agricultural Research, 15: 87-100.
R Core Team. R: A language and environment for statistical computing [Internet]. Vienna, Austria: R Foundation for Statistical Computing; [cited]. Available from: http://www.R-project.org/
Ragusa S. 1986. Five year study on population fluctuations of phytoseiid mites in a citrus orchard in Sicily. Acarologia, 27: 193-201.
Rasmy A.H., Zaher M., Albagoury M. 1972. Mites associated with citrus in the Nile Delta (UAR). Zeitschrift für angewandte Entomologie, 70: 183-186. doi:10.1111/j.1439-0418.1972.tb02168.x ![]()
Rosenheim J.A., Kaya H.K., Lester E.E., James J.M., Bruce A.J. 1995. Intraguild predation among biological-control agents: theory and evidence. Biological Control, 5: 303-335. doi:10.1006/bcon.1995.1038 ![]()
Sahraoui H., Tixier M.-S., Lebdi-Grissa K., Kreiter S. 2014. Diversity and abundance of Phytoseiidae (Acari: Mesostigmata) in three crop management strategies of citrus orchards in Tunisia. Acarologia, 54: 155-169. doi:10.1051/acarologia/20142123 ![]()
Satar S., Ada M., Kasap İ., Çobanoğlu S. 2013. Acarina fauna of citrus trees in eastern Mediterranean region of Turkey. IOBC-WPRS Bulletin, 95: 171-178.
Smith D., Papacek D.F. 1991. Studies of the predatory mite Amblyseius victoriensis (Acarina: Phytoseiidae) in citrus orchards in south-east Queensland: control of Tegolophus australis and Phyllocoptruta oleivora (Acarina: Eriophyidae), effect of pesticides, alternative host plants and augmentative release. Experimental and Applied Acarology, 12: 195-217. doi:10.1007/BF01193467 ![]()
Tixier M.-S. 2018. Predatory mites (Acari: Phytoseiidae) in agro-ecosystems and conservation biological control: a review and explorative approach for forecasting plant-predatory mite interactions and mite dispersal. Frontiers in Ecology and Evolution, 6: 192. doi:10.3389/fevo.2018.00192 ![]()
Tsagkarakis A.E., Emmanouel N.G., Panou H.N., Kapaxidi E.V., Papadoulis G.T. 2011. Composition and seasonal abundance of mites associated with citrus in Greece. International Journal of Acarology, 37: 252-259. doi:10.1080/01647954.2011.558853 ![]()
Urbaneja-Bernat P., Ibáñez-Gual V., Montserrat M., Aguilar-Fenollosa E., Jaques J.A. 2019. Can interactions among predators alter the natural regulation of an herbivore in a climate change scenario? The case of Tetranychus urticae and its predators in citrus. Journal of Pest Science, 92: 1149-1164. doi:10.1007/s10340-019-01114-8 ![]()
Vacante V., Gerson U. 2012. Integrated control of citrus pests in the Mediterranean region [Internet]. Bentham Science Publishers. doi:10.2174/97816080529431120101 ![]()
van der Hammen T., de Roos A.M., Sabelis M.W., Janssen A.J.O. 2010. Order of invasion affects the spatial distribution of a reciprocal intraguild predator. Oecologia, 163: 79-89. doi:10.1007/s00442-010-1575-7 ![]()
Warburg S., Inbar M., Gal S., Salomon M., Palevsky E., Sadeh A. 2019. The effects of a windborne pollen-provisioning cover crop on the phytoseiid community in citrus orchards in Israel. Pest Management Science, 75: 405-412. doi:10.1002/ps.5129 ![]()
Zeileis A. 2004. Econometric computing with HC and HAC covariance matrix estimators. Vienna: Department of Statistics and Mathematics, Wirtschaftsuniversität. doi:10.18637/jss.v011.i10 ![]()



2019-07-10
Date accepted:
2019-10-22
Date published:
2019-11-07
Edited by:
Tixier, Marie-Stéphane

This work is licensed under a Creative Commons Attribution 4.0 International License
2019 Warburg, Sharon; Gafni, Roni; Inbar, Moshe; Gal, Shira; Palevsky, Eric and Sadeh, Asaf
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)