




Supplementary description of Digamasellus punctum (Berlese) (Mesostigmata, Digamasellidae), and a key to the world species of Digamasellus Berlese
Hosseini, Leila1 ; Maroufpoor, Mostafa2 and Kazemi, Shahrooz3
1Department of Plant Protection, Faculty of Agriculture, University of Kurdistan, Sanandaj, Iran.
2Department of Plant Protection, Faculty of Agriculture, University of Kurdistan, Sanandaj, Iran.
3✉ Department of Biodiversity, Institute of Science and High Technology and Environmental Sciences, Graduate University of Advanced Technology, Kerman, Iran.
2019 - Volume: 59 Issue: 3 pages: 395-405
https://doi.org/10.24349/acarologia/20194341ZooBank LSID: 83FEF664-B73A-409D-93D4-28B68D1328D5
Original research
Keywords
Abstract
Introduction
Berlese (1905) proposed Digamasellus as a subgenus of Gamasellus Berlese, and designated a new species, G. (D.) perpusillus, as its type species. Lindquist (1975) reviewed conceptual and nomenclatural problems of Digamasellus and Dendrolaelaps Halbert, presented a complete diagnosis for these genera and also for the family Digamasellidae and showed that Digamasellus perpusillus is junior synonym of D. punctum (Berlese, 1904). Although before it, several species belonging to other genera had been described under Digamasellus by some authors (e.g. Womersley 1942; Leitner 1949; Ryke 1962; Hurlbutt, 1967), a few subsequent authors used the previous taxonomic rank and described/referred to some species belonging to other genera as members of Digamasellus (e.g. Bhattacharyya 1978; Pugh 1993; Castilho et al. 2016). Afterward, the only described species under Digamasellus that well match with all diagnostic characters of the genus is D. variabilis Wiśniewski & Hirschmann, 1989. Therfore, following Lindquist's (1975) diagnosis for Digamasellus, this genus currently comprises only three described species: D. punctum, D. australis Lindquist, 1975 and D. variabilis.
Mites of the genus Digamasellus usually occur in high organic substrates such as manure and rotting organic material and also under the bark of trees (Hirschmann 1960; Lindquist 1975, Wiśniewski & Hirschmann 1989). Hirschmann (1960) redescribed D. punctum and illustrated the dorsal idiosoma, epistome, part of leg II of female, and the ventral idiosoma and gnathosoma (partially) of deutonymph, male and female of the species. However, details of some structures such as the palp and the legs and also the peritrematal shield of the species have not been presented yet.
The Iranian mites of the family Digamasellidae are poorly known. Until now, only 12 species belonging to two genera of digamasellid mites have been recorded from Iran, including 11 Dendrolaelaps Halbert and one Dendroseius Karg species (Faraji et al. 2006; Kazemi & Rajaei 2013; Abolghasemi & Kazemi 2016; Nemati et al. 2018; Moradi Faradonbe et al. 2018). Haddad Irani-Nejad et al. (2001) reported the genus Digamasellus from Iran on the basis of a male specimen of an unknown species. Apparently, this specimen got lost (Haddad Irani-Nejad pers. comm. with SK), so we could not examine it. However, in spite of the poor-quality illustrations of the species (of the male specimen), the small anal opening of the species clearly indicates that it does not belong to the genus Digamasellus. Additionally, the presence of setae st5 on the posterior edges of the sternitigenital shield demonstrates that the species does not belong to the family Digamasellidae. So, the report of Digamasellus punctum from Iran in this paper effectively represents the first record of the genus from the country. Despite the relatively good redescription of D. punctum by Hirschmann (1960), he did not present a complete description/illustration of the legs, palp, peritreme and peritrematal shield and also idiosomal adenotaxy, poroidotaxy and sigillotaxy of the species. Therefore, here we present a complete description of the female of the species based on a single collected specimen in western Iran. Additionally, a key to the world species of Digamasellus is presented.
Material and methods
The mite specimen was removed from a sample of cow manure collected in Javanrood Region, Kermanshah Province by a Berlese-Tullgren funnel, cleared in Nesbitt's fluid and then mounted on a microscope slide using Hoyer's medium.
Morphological observations, measurements and illustrations were made using a compound microscope equipped with differential interference contrast and phase contrast optical systems, and a drawing tube (Olympus BX51). Measurements were made in micrometers (μm). The length of podonotal and opisthonotal shields was taken from the anterior to posterior shield margins along the midline; the length of the idiosoma was measured at the same level and the width at level of setae j6. The width of the sternal shield was taken from lateral margins of the shield at the level of setae st2, and its length from the level of poroids iv1 to posterior margin of shield along the midline. The length of epigynal shield was measured from the anterior margin of the hyaline extension to the posterior margin of the shield along the midline; shield width was taken at level of the genital setae. The ventrianal shield length and width were measured along the midline from the anterior to posterior margins, including the cribrum, and at the broadest level, respectively. The legs' lengths were measured from the base of the coxae to the apex of tarsi, excluding pretarsus. The length of the second cheliceral segment was taken from its base to the apex of the fixed digit. The length of the fixed cheliceral digit was measured from the dorsal lyrifissure to the apex and that of the movable digit from the base to apex.
Notations of idiosomal setae follow that of Lindquist & Evans (1965), and that for legs and palp setae follow that of Evans (1963, 1964). Notations of idiosomal pore-like structures as glandular openings (circular symbols) versus poroids (oval-shape symbols) follow mostly Athias-Henriot (1971, 1975), adapted by Kazemi et al. (2014).
Results
Digamasellus Berlese, 1905
Gamasellus (Digamasellus) Berlese, 1905.
Type species: Gamasellus (Digamasellus) perpusillus Berlese, 1905 (= Cyrtolaelaps punctum Berlese, 1904), by original designation.
Diagnosis
The genus diagnosis of Lindquist (1975) was followed.
Note — Lindquist (1975) indicated the absence of seta z3 in Digamasellus species. Setae z3 and s2 both appeared in the deutonymphal instar in Gamasina, but s2 is inserted more laterally in comparison with z3. Therefore, the seta located posterolaterad to z2 in Digamasellus species is named z3 (instead of s2) in this paper.
Lindquist (1975) referred to only two described species belonging to the genus Digamasellus (he had examined most related species in the Berlese collection in 1961 and 1971; pers. comm. with SK). However, recently Castilho et al. (2016) transferred some species described by Berlese (1920) under Gamasellodes (Digamasellus) to the genus Digamasellus, including G. ( D.) gracilis, G. ( D.) rhodacaroides and G. ( D.) simplex. Berlese's (1920) descriptions of these species are inadequate. Therefore, we tried to obtain additional information about these species based on photographs taken of the type material in the Berlese collection in Florence by the Curator (Roberto Nanneli). Based on the photographs of the type material of G. ( D.) gracilis and G. ( D.) rhodacaroides, although quality of the slides was not good, both of these species can be easily excluded from the genus Digamasellus by their small anal opening. Although Berlese (1920) described G. ( D.) simplex with a small anal shield bearing only circumanal setae that clearly indicates the species does not belong to Digamasellus (also mentioned by Ryke (1962)), no specimens of G. ( D.) simplex were available in the Berlese collection. Castagnoli and Pegazzano (1985) also pointed out that this species is indicated with an ''m'' in the ''Catalogue of the Berlese Acaroteca'' (page 467) referring to its absence (''missing''). Therefore, we considered it as nomen dubium. Berlese (1920) also described G. ( D.) reticulatus that based on the photographs of its type material (deutonymph), and although this species does not have incisions on anterior margin of the opisthonotal shield (as in Dendrolaelaps), it has a small anal opening which excluds it from Digamasellus. Leitner (1949) described Digamasellus fallax very briefly, but Shcherbak (1980) redescribed it as a member of the genus Dendrolaelaps and the exclusion of the species is confirmed here based on the following characters: (1) presence of four scleronoduli behind setae j5; (2) anal opening very small; and (3) presence of two incisions in anterior margin of the opisthonotal shield. Bhattacharyya (1978) described Digamasellus sitalaensis lacking information about leg chaetotaxy. However, it can be easily excluded from this genus by the following characters: (1) podonotal shield bears 22 pairs of setae, s2 and z3 both present; (2) the sternal shield has three pairs of setae (st1-st3), and setae st4 inserted off the shield; and (3) four scleronoduli present behind setae j5. To establish its correct taxonomic placement, the type specimens should be carefully examined, but it was not possible in this study.
Digamasellus punctum (Berlese, 1904)
Cyrtolaelaps punctum Berlese, 1904: 262.
Gamasellus (Digamasellus) perpusillus Berlese, 1905: 234.
Redescription
Female (Figures 1-5)
Idiosoma — Oval shape, 298 long, 205 wide.
Dorsal idiosoma — (Figures 1A-B). Podonotal shield length 151, width 184, shield surface mostly smooth, with a few submarginal longitudinal lines, bearing 21 pairs of setae (j1-j6, z1-z6, s1, s3-s6, r2-r5), five pairs of poroids and two pairs of gland pores (gd1, gd2); opisthonotal shield length 154, width 156, shield surface smooth, with 18 pairs of setae (J1-J5, Z1-Z5, S1-S5, R3-R5), 11 pairs of poroids and three pairs of gland pores (gd6, gd8-gd9), gd8-gd9 adjacent as enlarged double external pores (Figure 1A). Podonotal and opisthonotal shields setae mostly smooth, some of posterior setae on opisthonotal shield with a few minute barbs (e.g. J3-J4, Z2-Z4, S3-S5), dorsal setae usually with a small node basally, Z5 longest (58), apically slightly spatulate, J5 very short (8), blunt (Figure 1B), j1 and z1 short (9–10), smooth and needle like, rest of podonotal shield setae 14–20 long and those on opisthonotal shield 15–30 long. One unpaired seta R2 (12) asymmetrically present on left side in soft cuticle.
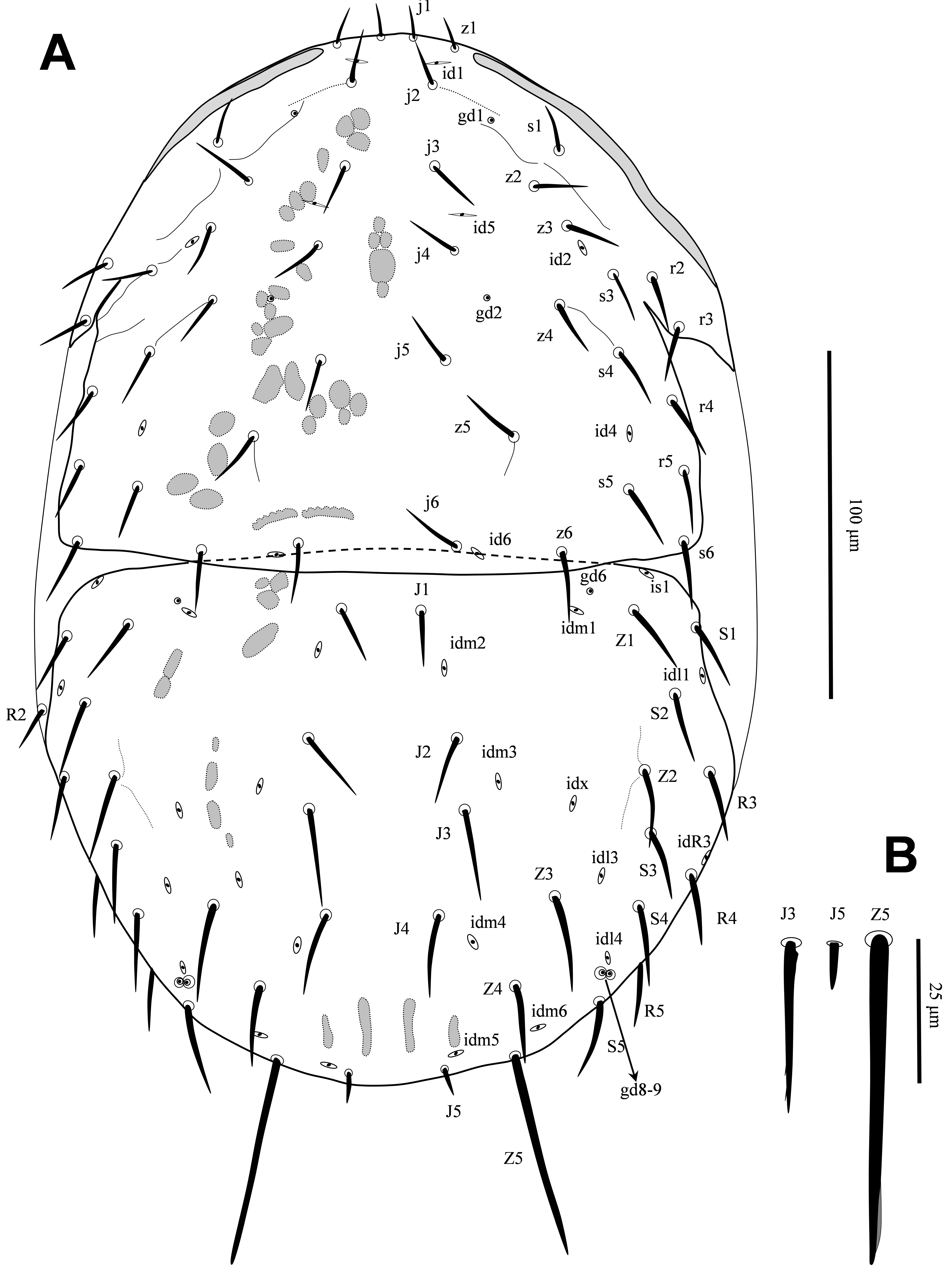
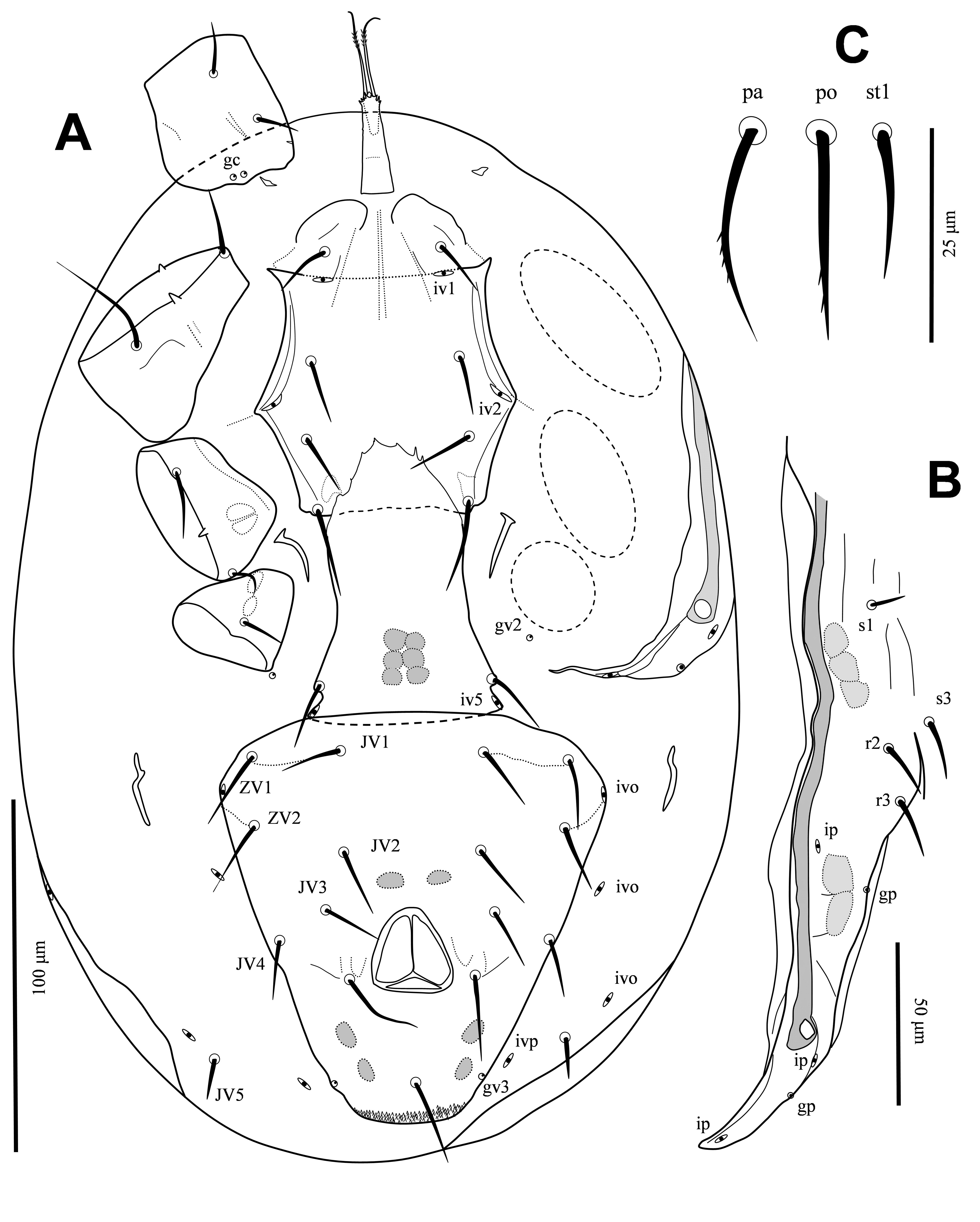
Ventral idiosoma — (Figures 2, 3A). Tritosternal base columnar and denticulate anteriorly, 26 long; laciniae free, 26-28 long, mostly smooth, with short dense barbs in subapical region (Figure 3A). Presternal region lightly sclerotized, bearing setae st1 (16). Anterior margin of sternal shield indistinct, shield surface smooth, 66 long, 63 wide; shield with three pairs of setae, st2 (18), st3 (20), st4 (27), and two pairs of distinct poroids, iv1-iv2. Endopodal platelets between coxae III-IV narrow, free. Epigynal shield 72 long, 53 wide, anterior hyaline margin of shield convex with few short teeth, posterior margin of shield slightly convex, shield surface smooth (except for median undersurface sigillar patterns), shield with short depressions immediately behind st5 (18 long) level, poroids iv5 on posterolateral edges of shield. Ventrianal shield subtriangular, 116 long, 110 wide, shield surface smooth, with six pairs of smooth pre-anal setae, JV1-JV4 (24–26), ZV1-ZV2 (26–28), circumanal setae with minute barbs (Figure 2C), postanal seta (26) slightly shorter than paranals (28), cribrum narrow; with one pair of poroids ivo antrolaterally on shield between ZV1-ZV2; gland pores gv3 at level of postanal seta on shield; anus large, subtriangular, 23 long, 24 wide; unsclerotized region of opisthogaster with gland pores gv2 behind coxae IV, a pair of narrow metapodal platelets, three pairs of poroids and setae JV5 (15) (Figure 2A). Peritrematal shields well developed, anteriorly fused to dorsal shield at mid-level of setae s3-s4, with three pairs of poroids and two pairs of gland pores; peritremes long, anteriorly reaching close to z1 (Figure 2B). Sperm-reception structures associated with coxae IV, but damaged and not clear after mounting.
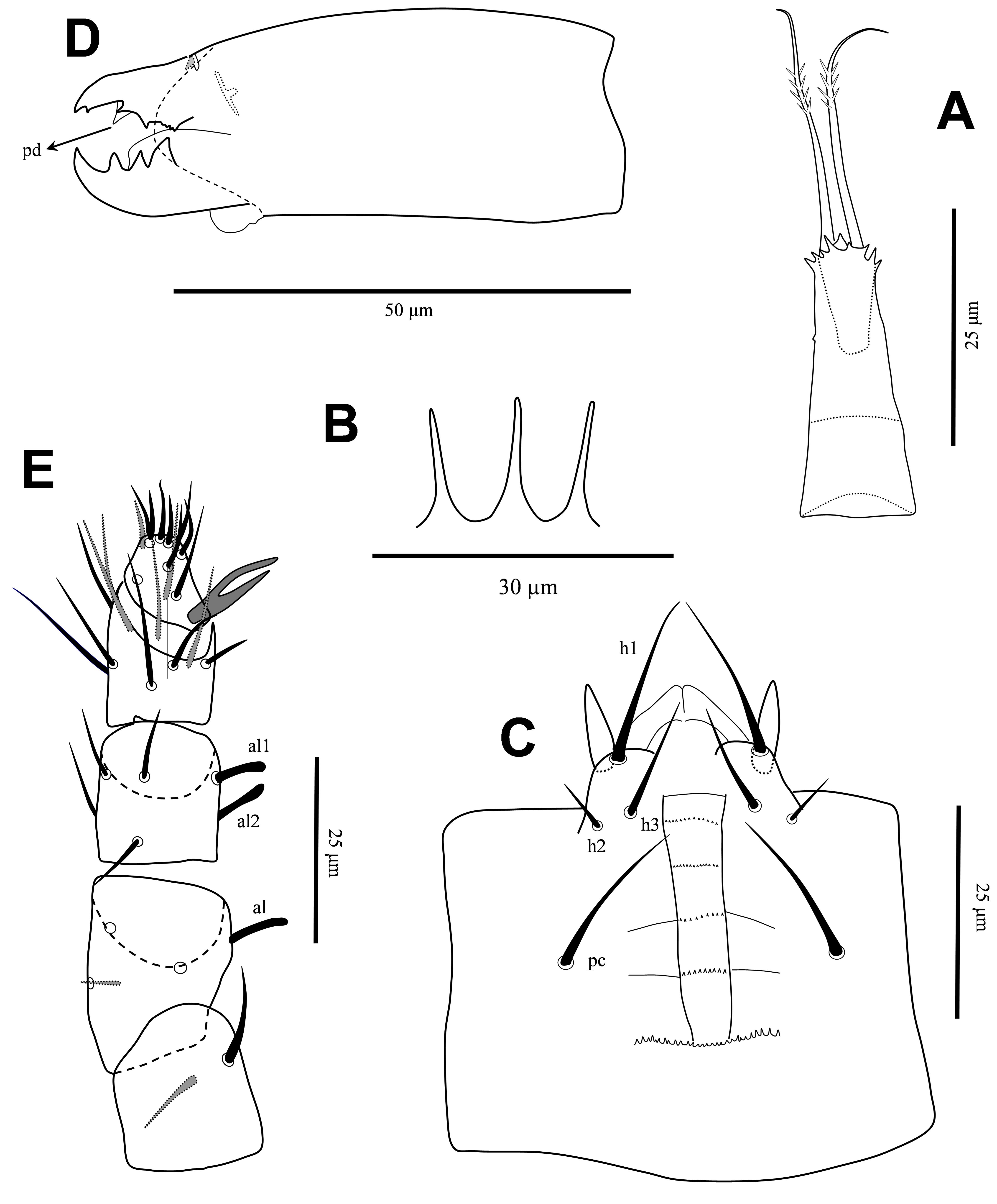
Gnathosoma — Epistome with three short, narrow projections (Figure 3B). Corniculi slender, straight, 11 long; internal malae shorter than corniculi; hypostomal setae smooth, h1 (20), h2 (7), h3 (14), pc (17); deutosternal groove relatively wide, with five rows delimited by lateral lines, anterior row smooth, others multidenticulate, with a wide basal denticulate row, almost three times broader than delimited rows (Figure 3C). Second segment of chelicera 61 long; movable digit 20 long, with three large teeth, basal tooth markedly larger; fixed digit 17 long, with a very short offset subapical tooth, two short median teeth in addition to few minute teeth basally, pilus dentilis lightly sclerotized, slightly widened at base (Figure 3D, arrow). Palp 88 long, setae v1-v2 on palptrochanter slightly thickened, setae al on palpfemur and setae al1-al2 on palpgenu spatulate, other palp setae narrow, without elongate setae on palptarsus; palptarsus apotele two tined (Figure 3E).
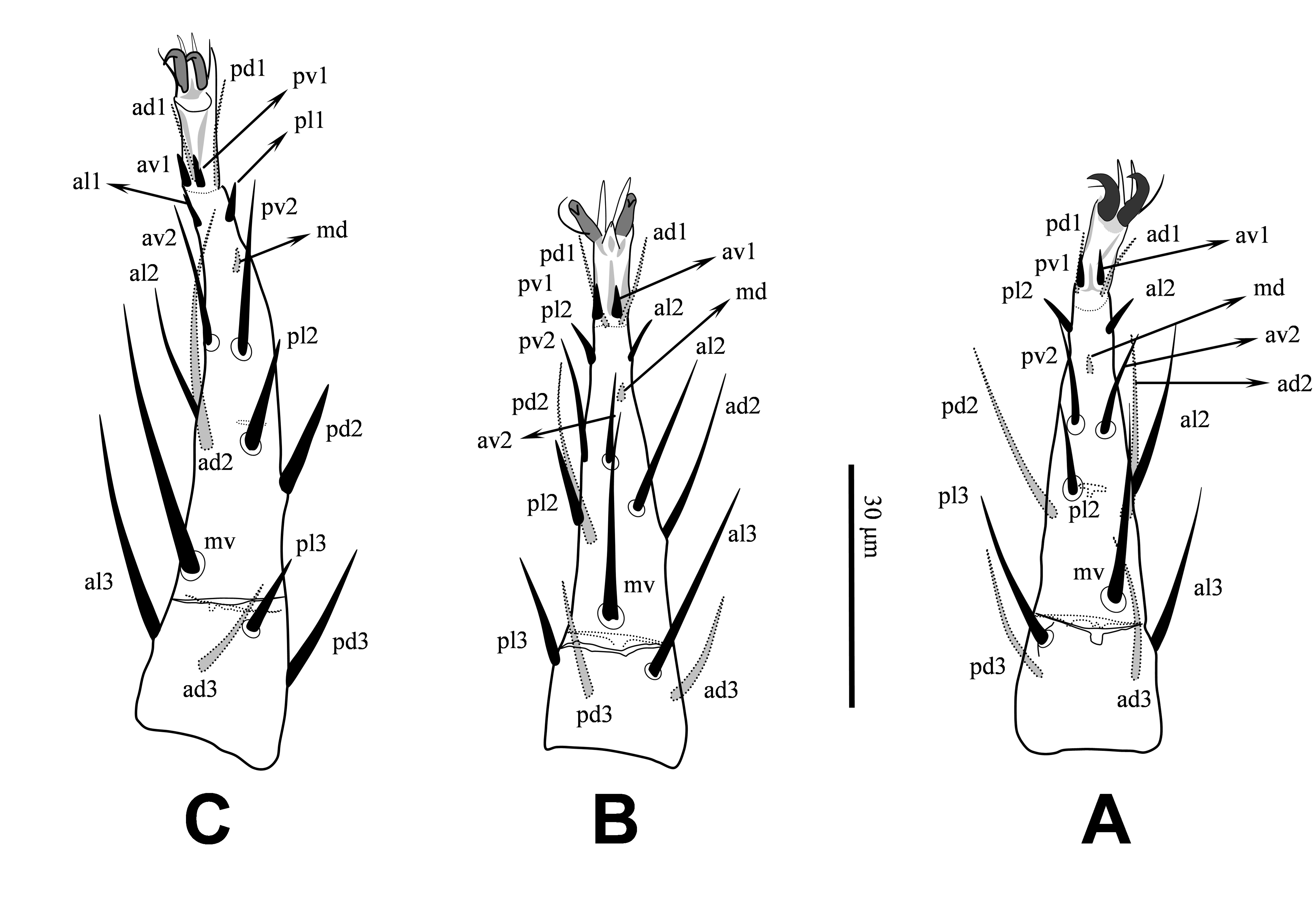
Legs — All legs with pretarsi and paired claws; pretarsi of legs II-IV each with one pair of relatively long pointed membranous processes arising from between bases of claws and extending well beyond tips of claws length (Figures 5A-C). Length of legs I-IV 252, 186, 177 and 215, respectively; length of pretarsi I-IV 19-22. Setation for legs I-IV, respectively, for trochanters 6 – 5 – 5 – 5; femora 13 (2 – 3/2, 2/2 – 2), 10 (2 – 2/2, 2/1 – 1), 6 (1 – 2/1, 2/0 – 0), 6 (1 – 2/1, 1/0 – 1); genua 12 (2 – 3/2, 2/1 – 2), 11 (2 – 3/1, 2/1 – 2), 8 (2 – 2/1, 2/0 – 1), 7 (1 – 2/1, 2/0 – 1); tibiae 12 (2 – 3/2, 2/1 – 2), 10 (2 – 2/1, 2/1 – 2), 8 (2 – 1/1, 2/1 – 1), 7 (1 – 1/1, 2/1 – 1) (Figures 4A-E); and tarsi II-IV each with 18 setae; all legs setae smooth, mostly short (Figures 5A-C).

Material examined — One adult female, collected in cow manure in western Iran, Javanrood Region, Kermanshah Province (34°48'15'' N; 46°38'49'' E), altitude 1437 m above sea level, 6 April 2018, coll. L. Hosseini, deposited in Acarological Collection, Institute of Science and High Technology and Environmental Sciences, Graduate University of Advanced Technology, Kerman, Iran.
Discussion
In Hirschmann's (1960) redescription of D. punctum, a seta inserted well behind the insertion of S2 was designated as R1. However, the normal position of R1 in Gamasina is more anterior, slightly behind S1 (Lindquist & Evans 1965). Lindquist (1975) demonstrated the absence of R1 in the protonymph, deutonymph and adults of Digamasellus and his hypothesis was confirmed by description of all postembryonic instars of D. variabilis by Wiśniewski & Hirschmann (1989). On the other hand, normal insertion of poroids idR3 is between setae R3 and R4, so it can be useful to identify these two setae too. Seta R2 is inserted in soft cuticle of D. punctum and D. variabilis, but it is on the opisthonotal shield in D. australis (Lindquist 1975; Wiśniewski & Hirschmann 1989).
One pair of ''double gland pores'' are distinctly present in D. punctum between setae S4-S5 (denoted as PS4 and PZ4 by Hirschmann (1960)) and also illustrated by Lindquist (1975) for D. australis in the same region. Wiśniewski & Hirschmann (1989) illustrated a pair of pore-like structures each including only one of these structures between the mentioned setae for D. variabilis, and no other pore-like structures on the dorsal shields of the latter in contrast to many pairs of pore-like structures shown by Hirschmann (1960) for D. punctum, 21 pairs by Lindquist (1975) including eight pairs on the podonotal shield (gv4 present near the base of seta s4) and 13 pairs on the opisthonotal shield for D. australis and also 22 pairs including seven pairs on the podonotal shield (gv4 absent) and 15 pairs on the opisthonotal shield in the current paper for D. punctum. On the other hand, in the original description of D. australis and D. variabilis only poroids iv1-iv2 were illustrated on the sternal shield of these species, but Hirschmann (1960) illustrated iv3 for D. punctum, although they were not present on the shield in the specimen examined by us and also in three females of this species deposited in the Canadian National Collection of Insects and Arachnids (CNCI), Science & Technology Branch, Agriculture & Agri-Food Canada, Ottawa (personal communication of one of the authors (SK) with E.E. Lindquist).
We also observed fungal spores inside the ventriculus of the collected mite that confirms fungivorous behavior of D. punctum as reported by Lindquist (1975).
Key to world species of Digamasellus (female)
1. Short post-stigmatic extension of peritreme present; coxa II with seta pv slightly shorter or longer than av (pv/av ≈ 1.3)
...... 2
— Short post-stigmatic extension of peritreme absent; coxa II with seta pv considerably longer than av (pv/av ≈ 2)
...... D. punctum
2. Seta Z5 slightly longer than J5 (Z5/J5 ≈ 1.6); ventrianal shield conspicuously wider than long; setae R2 inserted on opisthonotal shield
...... D. australis
— Seta Z5 considerably longer than J5 (Z5/J5 ≈ 6–7); ventrianal shield as long as wide; setae R2 inserted in soft cuticle
...... D. variabilis
Acknowledgments
We wish to thank Dr. Evert E. Lindquist for his valuable suggestions and also his help with some observations on female of Digamasellus punctum deposited in CNCI. We thank Roberto Nannelli for his valuable help in providing photographs from the type specimens of D. punctum, Gamasellus (Digamasellus) rhodacaroides, G. (Digamasellus) gracilis and G. (Digamasellus) reticulatus deposited in the Berlese collection, Florence, Italy and also information about deposition of G. (Digamasellus) simplex. We also thank Dr. Edward A. Ueckermann for his valuable suggestions.
References
Abolghasemi S., Kazemi, S. 2016. Report of a new species of Dendrolaelaps Halbert (Acari: Mesostigmata: Digamasellidae) for Iran. In: Talaei-Hassanloui R., Rahimi S., Ebrahimi V. (Eds). Proceedings of the 22th Iranian Plant Protection Congress, Karaj, Iran, p. 510.
Athias-Henriot C. 1971. La divergence néotaxique des Gamasides (Arachnides). Bull. Sc. Bourgogne, 28: 93-106.
Athias-Henriot C. 1975. Nouvelles notes sur les Amblyseiini II. Le relevé organotaxique de la face dorsale adulte (Gamasides Protoadenique, Phytoseiidae). Acarologia, 17: 20-29.
Berlese A. 1904. Acari nuovi. Manipulus II. Redia, 1: 258-280.
Berlese A. 1905. Acari nuovi. Materiali per "Manipulus V". Redia, 2: 231-238.
Berlese A. 1920. Centuria quinta di Acari nuovi. Redia, 14: 143-197.
Bhattacharyya S.K. 1978. Five new species and a new record of Mesostigmata (Acari) from West Bengal, India. Indian J. Acarol., 2(2): 78-87.
Castilho R.C., Silva E.S., Moraes G.J. de, Halliday B. 2016. Catalogue of the family Ologamasidae Ryke (Acari: Mesostigmata). Zootaxa, 4197 (1): 1-147. doi:10.11646/zootaxa.4197.1.1 ![]()
Castagnoli M., Pegazzano F. 1985. Catalogu of the Berlese Acaroteca. Istituto Sperimentale per la Zoologia Agraria, Firenze, Italy, 490 pp.
Evans G.O. 1963. Observations on the chaetotaxy of the legs in the free-living Gamasina (Acari: Mesostigmata). Bull. Br. Mus. (Nat. Hist.), Zool., 10: 275-303. doi:10.5962/bhl.part.20528 ![]()
Evans G.O. 1964. Some observations on the chaetotaxy of the pedipalps in the Mesostigmata (Acari). Ann. Mag. Nat. Hist. (Ser. 13), 6: 513-527. doi:10.1080/00222936308651393 ![]()
Faraji F., Sakenin-Chelav H., Karg W. 2006. A new species of Dendroseius Karg from Iran (Acari: Rhodacaridae), with a key to the known species. Zootaxa, 1221: 63-68. doi:10.11646/zootaxa.3768.1.3 ![]()
Haddad Irani-Nejad K., Kamali K., Maleki Milani H. 2001. Mites of Digamasellidae, Rhodacaridae, Phytoseiidae and Ascidae of the cotton fields in Moghan Plain. J. Agricul. Sci. & Technol. (15)1: 23-33.
Hirschmann W. 1960. Gangsystematik der Parasitiformes. 3. Die Gattung Dendrolaelaps Halbert 1915. Acarol. Schrift. Vergl. Milbenk., 3: 1-27, pls. 1-28.
Hurlbutt H.W. 1967. Digamasellid mites associated with bark beetles and litter in North America. Acarologia, 9: 479-534.
Kazemi S., Rajaei A. 2013. An annotated checklist of Iranian Mesostigmata (Acari), excluding the family Phytoseiidae. Persian J. Acarol., 2(1): 63-158. doi:10.22073/pja.v2i1.9950 ![]()
Kazemi S., Rajaei A., Beaulieu F. 2014. Two new species of Gaeolaelaps (Acari: Mesostigmata: Laelapidae) from Iran, with a revised generic concept and notes on significant morphologi¬cal characters in the genus. Zootaxa, 3861(6): 501-530. doi:10.11646/zootaxa.3861.6.1 ![]()
Leitner E. 1949. Zur Kenntnis der Gattung Digamasellus Berlese 1905. Zentbl. Gesamtgeb. Ent., 3: 51-62.
Lindquist E.E. 1975. Digamasellus Berlese, 1905, and Dendrolaelaps Halbert, 1915, with descriptions of new taxa of Digamasellidae (Acarina: Mesostigmata). Canadian Entomol., 107(1): 1-43. doi:10.4039/Ent1071-1 ![]()
Lindquist E.E., Evans G.O. 1965. Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina (Acarina: Mesostigmata). Mem. Entomol. Soc. Canada, 47: 1-64. doi:10.4039/entm9747fv ![]()
Moradi Faradonbe M., Khajehali J., Amani M., Moraza M.L. 2018. First report of two species of Digamasellidae (Acari: Mesostigmata) from Iran. In: Pourrahim R. (Ed). Proceedings of the 23th Iranian Plant Protection Congress, Gorgan, Iran, p. 1632.
Nemati A., Riahi E., Khalili-Moghadam A., Gwiazdowicz D.J. 2018. A catalogue of the Iranian Mesostigmata (Acari): additions and updates of the previous catalogue. Persian J. Acarol., 7(2): 115-191. doi:10.22073/pja.v7i2.36985 ![]()
Pugh P.J.A. 1993. A synonymic catalogue of the Acari from Antarctica, the sub-Antarctic Islands and the Southern Ocean. J. Nat. Hist., 27(2): 323-421. doi:10.1080/00222939300770171 ![]()
Ryke P.A.J. 1962. The subgenera Digamasellus Berl. And Euryparasitus Oudemans of the genus Cyrtolaelaps Berlese (Acarina: Rhodacaridae). Entomol. Soc. South. Africa, 25: 88-115.
Shcherbak, G.I. (1980) The Palearctic Mites of the family Rhodacaridae. Naukova Durnka, Kiev, 215 pp.
Wiśniewski J., Hirschmann W. 1989. Gangsystematik der Parasitiformes teil 510 neue Digamasellus- und Dendrolaelaps-arten (Trichopygidiina) aus Kuba. Acarologia, 30(4): 305-323.
Womersley H. 1942. Additions to the Acarina − Parasitoidea of Australia. Part I. T. Roy. Soc. South Aust., 66: 142-171.



2019-02-14
Date accepted:
2019-10-04
Date published:
2019-10-07
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2019 Hosseini, Leila; Maroufpoor, Mostafa and Kazemi, Shahrooz
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)