




Oribatid mites (Acarina, Oribatida) from French Guyana II. Two new species from superfamily Crotonioidea Thorell, 1876
Miko, Ladislav1
1Charles University Prague, Institute for Environmental Studies, Czech Republic.
2019 - Volume: 59 Issue: 3 pages: 348-363
https://doi.org/10.24349/acarologia/20194336ZooBank LSID: 8100ECA7-4A71-46A4-94AE-1A5E812B3C3B
Original research
Keywords
Abstract
Introduction
The oribatid fauna of French Guyana was completely unknown, with no published information before my previous work referring to mites from the family Rhynchoribatidae (Miko, 2016). In this second contribution on oribatids of French Guyana, the aim is to describe two new species from the superfamily Crotonioidea Thorell, 1876 Malaconothrus kawensis n. sp. and Cyrthermannia bifurcata n. sp., found in material collected in Kaw mountains (Montagne de Kaw) and in the vicinity of Saül, respectively, and to analyse their relationships within their relevant genera.
Methods
Material studied for this publication was collected in the range of Kaw Mountains (Malaconothrus kawensis n. sp.) and in the vicinity of Saül (Cyrthermannia bifurcata n. sp.). Samples were collected in tropical forest, and comprised mostly of decomposing litter and decaying wood, including mosses and lichens growing on the ground, on rocks and as epiphytes. Detailed information about samples, their composition, location and date of collection is provided in respective species descriptions. Samples were transported in plastic bags and extracted immediately for 3 days in field extractors, based on the principle of Berlese apparatus or entomological xero-photo-eclector and constructed from simple linen cloth with a small plastic funnel at the bottom. Electric bulb was used as a source of light and heat as much as the electricity supply allowed, and the rest of the time samples were left to dry in the sun as much as possible. Extracted material was collected into 75% alcohol (ethanol). Samples were transported to Europe and sorted. Individuals were studied in temporary open slides after maceration in lactic acid. Material after study is kept in 75% alcohol (ethanol) and held either in collections of Senckenberg Museum in Goerlitz (Germany), or in the private collection of the author (for details see description of individual species).
For description and detailed study, standard terminology summarised by Norton and Behan Pelletier (2009) was used. All measurements are given in micrometers. Body length is given as maximum length (from tip of the rostrum to posterior end of notogaster) and as a ventral length (from tip of the rostrum to posterior margin of ventral plate). Length of prodorsum was measured in dorsal view as a distance between tip of the rostrum and anterior margin of the notogaster. Maximum width of body was measured in the broadest part of the notogaster, usually between tips or slightly behind discidia, depending on the species. Maximum width of prodorsum was measured in the broadest part between lateral margins of pedotecta I.
Genus Malaconothrus Berlese, 1904
Genus has global distribution, with 84 known species (Subías, 2004, 2018). Two more species were described recently from Africa (Ermilov and Starý, 2019). The concept and generic definitions within the family Malaconothridae have been reviewed for several times (Weigmann, 1997, 2006; Subías, 2018; Colloff, 2013; Colloff and Cameron, 2013). As a result, the new definition of the genus was proposed (Colloff and Cameron, 2013) and species have been grouped into 5 quite well defined species groups: plumosus, opisthoseta, crispus, monodactylus and marginatus group (Colloff, 2013). The species found in the Kaw mountains clearly belongs to the ``plumosus" group, which is defined by Colloff (2013) as \\
``... monodactylous species with broad, well-developed notogastral ridges; a broad, sub-rectangular rostrum; barbed or smooth, thick rostral setae; five pairs of genital setae, often thick and dilated basally, with g5 considerably longer than the others and further from g4 than the distance between g4 and g3. The interlamellar seta is often very short, sub-equal in length to the exobothridial seta or only twice as long". Nevertheless, none of the 5 species known from Central and South America is similar to the new species. The only similar species are known from Japan (Yamamoto 1996; Yamamoto and Aoki, 1997), and they belong to the same subgroup according to Colloff (2013). Recently, Ermilov and Starý (2019) described two species from Cameroon with very similar general appearance to the new species. However, Malaconothrus found in French Guyana clearly differs from all these species and is described below as a new species. The differences to similar species are discussed in discussion and in attached determination key.
Malaconothrus kawensis n. sp.
ZOOBANK: F2AA5098-43CB-48EF-ADD7-DFBAD9A80483 ![]()
Figures 1, 2

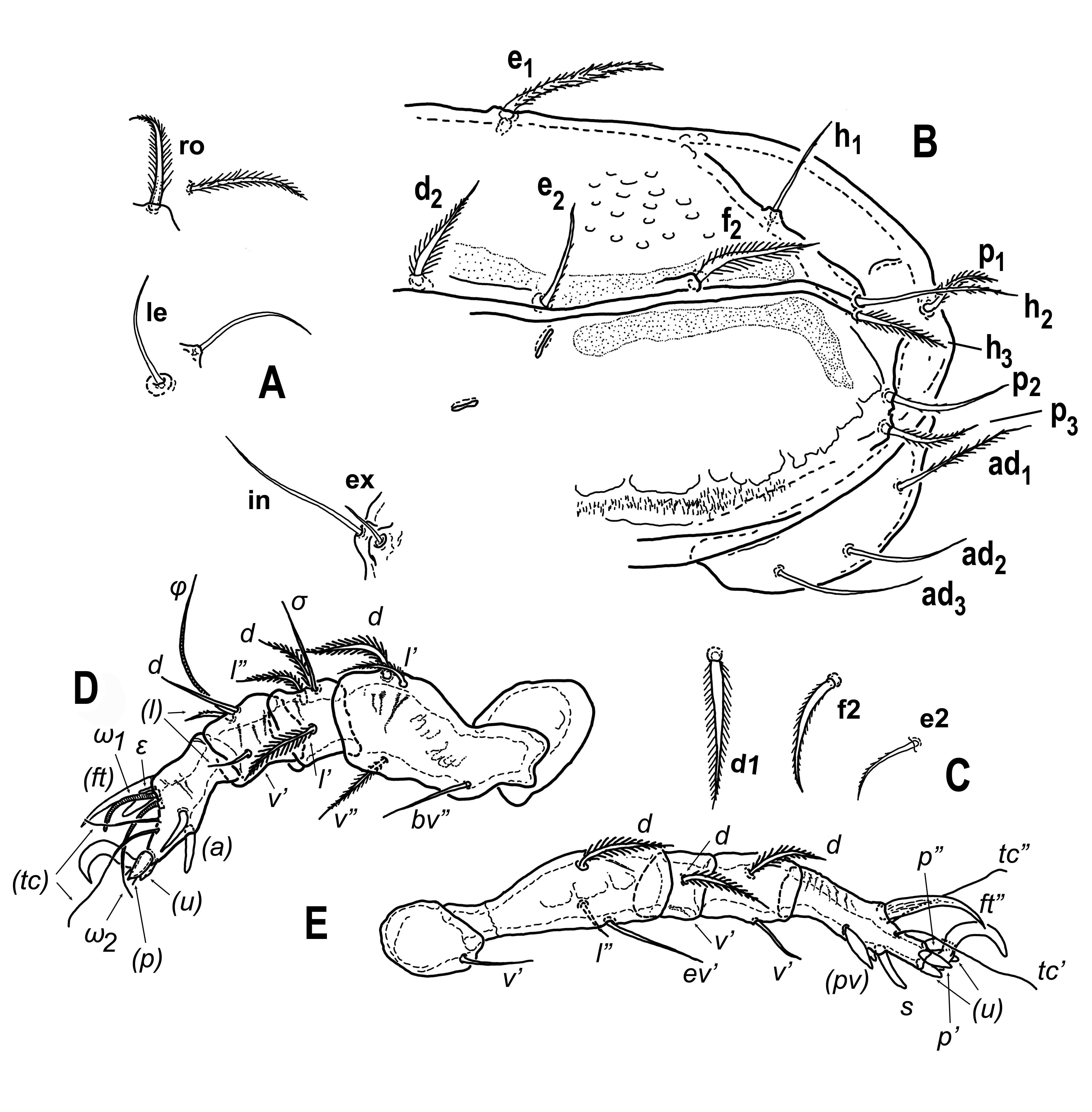
Diagnosis — Malaconothrus species from ``plumosus" group (Colloff, 2013), with most of notogastral setae slightly dilated and bilaterally barbed, and some notogastral setae (e2 , h1 , h2 , p2 ) smooth, setae h2 being the longest. Centrodorsal setae of notogaster d1 reaching significantly behind half distance of d1 and e1 . Prodorsal setae, particularly in, quite short, smooth, with only rostral setae densely bilaterally barbed. Epimeral setae 4b longest of epimeral setae IV and not positioned on posterior tectum of epimere IV. Dotted/porose areas present parallel to insertions of centrodorsal setae, also along lateral margins of notogaster and parallel to lateral edges of anogenital plate.
Material examined — 5 females collected 10-III-2012 in Kaw Mountains, in litter, dead leaves and rotting timber on the soil surface in tropical rainforest near to Coq du Roche Reserve, 4°34'2'' N, 52°11'58'' W (sample nr. LM-2012-008), French Guyana. Type and one paratype preserved in acarological collection of Senckenberg Museum in Goerlitz, 3 paratypes (one damaged) in collection of the author.
Description — Measurements – Values given for holotype, with range observed in paratypes in parentheses. Total body length 371 (365–385), prodorsum length 132 (130–136), prodorsum width 127 (125–132), width of notogaster 142 (142–145).
Integument – (Figs 1A-B, 2B). Body color pale, yellowish or white. Body suface with different types of sculpture: notogaster with small foveolae (diameter mostly around 2.5–3, up to 5 in posterior part of notogaster), with mutual distances larger than their diameter (5–6.5). Prodorsum with dense punctuation, in the area behind setae le with perforated cuticle (``chagrenation"), with maculae visible laterally and also between setae in. Ventrally densely punctate on epimeres, these also with muscle maculae. On both dorsal and ventral side of body, distinct areas of dense punctuation present, resembling porose areas of higher oribatids, some of these areas more distinctly framed, some not. These areas aligned antiaxially along insertions of setae e1 and f1 , axially to lateral setae (from c2 to the level of h1 insertion). Irregular areas also present along notogastral posterior ridges, and V-shaped areas present behind insertions of setae c1 . Similar, longitudinal areas present on ventral side, laterally along anogenital plate. Genital plates with more or less distinct, rugose surface sculpture.
Prodorsum – (Figs 1A, 2A). Rostrum conical, with very blunt tip or almost rounded. Short rostral carinae (roc) present, running posteriad from ro insertions, their anterior ends connected with distinct transversal line; this structure giving rostrum angular appearance in slightly posterodorsal view. Lateral carinae (lac) poorly developed, visible only in proximal part, not reaching insertions of setae le. Posterior of prodorsum behind insertions of setae in and ex narrowed, creating elbow-like postbothridial carinae. Indistinct transverse line visible between insertions of in, probably representing slightly thickened cuticle on anterior edge of deepened posterior part of prodorsum. Rostral setae (ro) medium long (31–39), slightly thickened at base, distinctly barbed. Lamellar setae (le) of about the same length as ro (32–39), simple setiform, smooth. Interlamellar setae (in) largest of prodorsal setae (47–50), smooth, setiform. Setae ex1 short, between one quarter and one third of in length (14–15), smooth, setiform. Setae ex2 absent.
Notogaster – (Figs 1A, 2B, 2C). Anterior margin straight, posterior margin slightly angulate in area of p1 insertions. Notogaster slightly dilated posteriad, broadest part is at level of setae f2 . Notogastral ridges present, well visible only in posterior part as inverted ``W" with anterior ridges reaching more or less posterior to e1 insertions. Distinct transverse ridge present between insertions of setae p1 in posterior part of notogaster. Setae c2 inserted on blunt tubercular thickening of cuticle. Notogastral setae of different length and form, d1 , e1 and h2 longest (42–49), c2 , c3 , e2 , h1 , h3 , p1 and p3 shortest (24–28). All notogastral setae barbed, most of them slightly thickened and with distinct barbs longer than width of setae, only e1 , h1 , p2 and h2 appearing almost smooth, but in detailed view each with at least few minute barbs. All lyrifissures distinct.
Ventral characters – (Fig. 1B). Gnathosoma as in other species in genus (Colloff and Cameron, 2013), setae a, m and h short, simple setiform. Epimeral region with distinct apodemes, epimeres IV with blunt posterior tectum projecting posteriad over anterior part of anogenital plate. Epimeral setal formula 3-1-3-3, epimeral setae mostly simple setiform and smooth (with only few minute barbs at most), except setae 3b and 3c which are distinctly barbed. Length of epimeral setae very different, 1a, 1c, 2a and 3a minute or vestigial, 4a and 4c short (around 7–8) and 1b, 3b, 3c and 4b somewhat longer (13–18). Anogenital plate with more or less parallel sides, length of genital plates 70, width 29, anal and adanal plates slightly longer (83), maximum width of anal plates 8, adanal 31. Five pairs of smooth genital setae present, with growing size from anterior to posterior, g1 (10) more than 2 times shorter than g5 (23). Alveoli of single pair of anal setae indistinct. Three pairs of quite long (38–44) adanal setae present, ad1 distinctly barbed, ad2 almost smooth or with some barbs, ad3 setiform and smooth. Adanal lyrifissures large, distinct, mostly transversal, located anteriorly to ad3 . Lyrifissures ips quite large, positioned near lateral edge of anogenital plate at level of anterior margin of anal plates.
Legs – (Figs 2D, 2E). Robust and rather short, as typical for genus, with smooth claws. Setae differently shaped, mostly (except tarsi) setifom or slightly thickened and barbed, on tarsi some setae strong, spiniform, with blunt or pointed and curved tip. Trochanteral setal formula 0-0-2-1, setae of trochanter III barbed. Only leg I and IV studied in detail, setal formula of leg I 0-4-3[1]-4(1)-11, setal formula of leg IV 1-3-2-2-10. Setae d of femur, genu and tibia I and IV , lateral setae of genu I as well as setae l' and v'' of femur I barbed similar to notogastral setae. Setae tc of tarsus on both leg I and IV long (44–46), filiform, tactile. Setae u, pv and ft' of leg I and p, u, s and pv of leg IV thick, spiniform, rather short and more or less blunt, seta ft'' of tarsus IV long (33), thickened, curved and acute, claw-like. Solenidia longer than setae d on genu and tibia of leg I (solenidion φ the longest, 47), tarsal solenidia as long as seta ft'' or longer, but shorter than setae tc. Famulus short, baculiform.
Derivatio nominis — The name kawensis relates to the typical locality, where the species was found – Montagne de Kaw.
Remarks — The species of genus Malaconothrus as redefined by Colloff and Cameron (2013) have been recently organised into 5 distinct species-groups (Colloff, 2013). Malaconothrus kawensis belongs clearly to ``plumosus" species group, originally proposed by Mahunka (1992) and extended by P. Balogh (1997). This group contains 28 known species with broad pan-tropical and subtropical distribution. The species of this group may be further sub-divided into 4 well defined groups, regarding development of notogastral setae (Colloff, pers. comm.): with notogastral setae all smooth (15 species); all barbed (9 species); notogastral setae e2 , some of h series, and p2 smooth, others barbed (3 species); and setae of c and p series and h3 smooth, others barbed (1 species). The new species described here belongs to the rather small subgroup of the ``plumosus" group, characterised by notogastral setae partly smooth (setae e2 , some of h series, p2 ) and partly slightly dilated and barbed (remaining setae). Three other species belong to this subgroup: M. kiiensis Yamamoto 1996, M. iriomotensis Yamamoto and Aoki, 1997 and M. variosetosus Hammer, 1971. Malaconothrus variosetosus can be easily distinguished by the different number of genital setae (6, vs. 5 in all other species including M. kawensis). In comparison to M. kawensis, it has much shorter notogastral setae, particularly c1 , d1 and e1 , which do not reach even half the distance to the following setae (vs. almost reaching it in M. kawensis), and barbed notogastral (and rostral) setae barbed only unilaterally (vs. bilateral in M. kawensis). The other two species of the subgroup appear to be more closely reated to M. kawensis – apart of mentioned characters, they also share weak development of lamellar ridges, presence of rostral carinae, presence of indistinct ridge between insertions of interlamellar setae and presence of broad, dotted/porose areas in position of the notogastral longitudinal ridges. These species also share presence of 5 genital setae, mostly concentrated in anterior half of the genital plates. Interesting similarity was found also on shape of adanal setae – in all three species they are longest of the ventral setae, and they may be variable regarding presence of bristles: Yamamoto and Aoki (1997) found that seta ad1 may be smooth or with bristles, similar to seta ad2 of the new species. The Japanese species M. kiiensis and M. iriomotensis, however, share some characters which are not present in the new species: longer prodorsal setae (in particular in, but also le and ro), relatively shorter notogastral setae (particularly centrodorsal, setae d1 reaching only to half distance between d1 and e1 ) with shorter and less dense bristles, and position of very short epimeral seta 4b (indicated in original descriptions of both species as 4a) on posterior tip of epimeral tectum IV. Malaconothrus kawensis has prodorsal setae shorter, centrodorsal setae of notogaster longer (d1 reaching far posterior of half distance d1 and e1 ) and with much denser and longer bristles, and epimeral setae 4b much longer and positioned much more anteriad, not on posterior tectum of epimere IV. The new species differs also by presence of V-shaped dotted/porose areas posterior to insertions of c1 , parallel along the notogaster laterally, and lateral to anogenital plate ventrally. Sculpture of notogastral cuticle is also different, M. kawensis has the notogaster with distinct and mutually quite distant small foveolae, the two Japanese species have notogaster with much finer ornamentation: very small and quite dense round foveolae or dots in M.kiiensis and dense punctuation in M. iriomotensis.
Two recently (Ermilov and Starý, 2019) described species from Cameroon – Malaconothrus crassilamellaris Ermilov and Starý, 2019 and Malaconothrus ciliarostralis Ermilov and Starý, 2019 have very similar general appearance to the new species (particularly very similarly developed notogaster, with similar porose areas along notogastral ridges, and similar length of notogastral setae). M. ciliarostralis has further similarities in development of some setae (barbed rostral setae, most of notogastral setae bilaterally barbed). Both species from Cameroon are easily distinguished from the new species, as the notogastral setae are either all smooth (M. crassilamellaris) or all barbed (M. ciliarostralis), and the development of ventral setation differs as well – particularly seta 4b (designated in paper of Ermilov and Starý as 4a) is long and not positioned on posterior tectum of epimere IV in M. kawensis, unlike in both Cameroon species (and two Japanese species discussed above).
In the following determination key, the main differences between species of ``plumosus" group from the closely related subgroup of species discussed above (indicated in key as (a) following the species name) as well as species from other subgroups of ``plumosus" group known from Latin America (indicated as (b) following the species name). Remaining species of the group are presented only as overal numbers in subgroups, not further elaborated.
\subsubsection*{Key to the species of the genus Malaconothrus } (plumosus group)
1. All notogastral setae smooth (15 species overall, 2 in Latin America)
...... 3
— At least some notogastral setae with sparse or dense, unilateral or bilateral bristles
...... 2
2. All notogastral setae barbed, with bristles (9 species overall, 2 in Latin America)
...... 4
— Only some notogastral setae barbed, at least one seta from series p or h smooth
...... 5
3. Rostral setae thickened, lamellar and adanal setae simple setiform. Adanal setae and setae of p series subequal in length. Genital seta g5 not more than 2-times longer than g4 . Size 410
...... Malaconothrus robustus Hammer, 1958 (b)
— Rostral setae smooth setiform, lamellar and adanal setae slightly widened, narrowly blade-shaped. Lamellar setae inserted on distinct tubercles. Setae p1 and p2 very long, 1.5–2 times longer than adanal setae. Genital seta g5 about 4-times longer than g4 . Size 410
...... Malaconothrus rohri P. Balogh, 1997 (b)
4. Setae in very long, simple setiform, much longer than seta ex. All other prodorsal and all notogastral setae long (reaching or almost reaching insertion points of subsequent row), robust, blade-like, with short, sparse barbs, adanal and genital setae also broadened, blade-like. Genital plates with 5 genital setae. Size 417–447
...... Malaconothrus hauseri Mahunka, 1984 (b)
— Setae in short, simple setiform,subequal in length to ex. Setae le and ro much longer, robust, bilaterally barbed. All notogastral setae short (not reaching even half of the distance to insertions of next row), setiform, densely bilaterally barbed. Ventral setae all also heavily barbed. Genital plates with 4 genital setae. Size 300–315
...... Malaconothrus neoplumosus Balogh and Mahunka, 1969 (b)
5. Only seta h1 of notogaster smooth, all other notogastral setae robust, mostly slightly thickened basally, bilaterally barbed. Rostral, lamellar and adanal setae also bilaterally with short barbs. Size 375
...... Malaconothrus granulosus Palacios-Vargas and Iglesias, 1997 (b)
— 3 or more notogastral setae smooth, remaining setae of different shape, barbed bi- or unilaterally, with barbs of different length
...... 6
6. Notogastral setae relatively short, robust, more or less broadened. Setae of c series, p series and h3 always smooth, at least some of others unilaterally, finely barbed
...... (2 species: Malaconothrus keriensis Hammer, 1966 with 6 genital setae and Malaconothrus kotozenus Fujikawa, 2005 with 5 genital setae, none of them in Latin America)
— Longer notogastral setae (e2 , some of h series, p2 ) smooth, all others slightly thickened and bilaterally barbed
...... 7
7. Prodorsal setae, particularly setae in short, centrodorsal setae of notogaster longer (d1 reaching far posterior of half distance d1 and e1 ) and with much denser and longer bristles. Epimeral setae 4b longest of epimeral setae IV and positioned much more anteriorly, not on posterior tectum of epimere IV. Dotted/porose areas present along lateral margins of notogaster and parallel to lateral edges of anogenital plate. Size 365–385
...... Malaconothrus kawensis n. sp. (a, b)
— Prodorsal setae, particularly in quite long, overhanging lateral edges of prodorsum. Centrodorsal setae d1 of notogaster reaching only to half distance between d1 and e1 , or less. Epimeral setae 4b very short, positioned on posterior projection of tectum of epimere IV. Dotted/porose areas along lateral margins of notogaster and parallel to lateral edges of anogenital plate absent
...... 8
8. Rostral setae strong, densely barbed, curled. Lamellar setae straight and rather long, bilaterally barbed. Pedotectum I angular, short humeral ridges present on notogaster between setae c2 and cp. Notogastral seta e2 significantly longer than surrounding notogastral setae d2 and f2 . Setae p2 and p3 subequal in length, both barbed. Genital setae long, seta g5 reaching almost to the posterior end of genital plate. Anal setae absent (only alveoli present) Size 405–470
...... Malaconothrus kiiensis Yamamoto, 1996 (a)
— Rostral and lamellar setae smooth, setiform, curved backwards. Pedotectum I obtuse, humeral ridges absent or indistinct. Notogastral seta e2 subequal in length to surrounding notogastral setae d2 and f2. Setae p2 smooth, much longer than barbed p3 . Genital setae short, seta g5 not reaching the posterior end of genital plate (distance to it is greater than length of seta g5 ). Short anal setae present. Size 385–435
...... Malaconothrus iriomotensis Yamamoto and Aoki, 1997 (a)
Genus Cyrthermannia Balogh, 1958
Balogh (1958) proposed the genus Cyrthermannia for Nanhermanniidae with 6–8 large, blunt and rounded protuberances on posterior part of the notogaster; and notogastral setae much shorter than those of Nanhermannia, and narrowly lanceolate. This definition is very broad and simple, but genus as such was established and accepted by most authors. A more modern diagnosis of the genus was never given. Some additional characters of the genus may be extracted from later published determination keys. Balogh (1972) added the following diagnostic characters: notogaster without pteromorphs, areae porosae, sacculi or pori (Apterogasterina-Gymnonota); more than 6 pairs of genital setae (Polytricha), notogaster with 16 pairs of setae (!), concentric or excentric exuviae never present on notogaster, diagastric (with semicircular, medially interrupted suture between genital and anal plates – Nanhermannioidea), notogastral setae simple, whole, without flagelliform apical section, short; posterior portion of notogaster with tubercles. Later, Balogh and Balogh (1992) placed the family within nothroid genera, and precised information about genus as follows: sensillus present, legs monodactyle, 9g, 2ag, 2an, 3ad, epimeral setal formula 3-1-3-3. Genital and anal plates of brachypyline type, separated by ventral plate. Anal plates with 2 pairs of setae. Semicircular, medially interrupted suture between genital and anal plates present (Nanhermanniidae). Notogaster with blunt tubercles bearing part of notogastral setae. Notogastral setae short, setiform or leaf-shaped, never bifurcate. Posterior protuberance of prodorsum with one triangular process on each side. Later, Hammer (1979) proposed a separate genus Bicyrthermannia for species differing from Cyrthermannia by having two distinct projections on the posterior side of interbothridial protuberances, and 16 setae on notogaster (even if her own figure shows only 15 notogastral setae). Later described species of both Cyrthermannia and Bicyrthermannia showed a variability in setation, particularly of genital plates (7–9 pairs of setae), but also on epimeres (with formulas 3-2-3-3, 3-1-3-4 and 3-2-3-4 also present). Variable number of notogastral setae was reported (from 13 to 16), but from descriptions it is difficult to judge if all setae were really counted. The figures sometimes indicate the possibility that some setae may have been omitted. In any case, the most common number of notogastral setae is 15 (and not 16 as given by Balogh, 1972). Given the presence of 15 notogastral setae in several species of Bicyrthermannia, the presence of two large spiniform tubercles on the posterior part of prodorsum against a single one in Cyrthermannia remains the only distinguishing character of the genus. Characteristics of legs are usually not given at all in most of described species, or only very scarce and general information is available. More detailed description of legs is available only for single species of Bicyrthermannia (Badejo and al., 2002).
Cyrthermannia bifurcata n. sp.
ZOOBANK: D343694D-0938-4F8B-9B7F-2CC6D457E0C0 ![]()
Figures 3, 4


Diagnosis — Cyrthermannia with short, smooth and distally bifurcate sensillus, with rostral protuberance and lamellar ridge present. Notogaster with distinct protuberances pronounced in both posterior and lateral parts of notogaster. Notogastral setae simple, setiform, smooth, some with short spur basally. Epimeral setae short setiform, 9 pairs of genital setae, 2 pairs of short and simple aggenital setae, 3 pairs of adanal setae which are the longest setae on ventral side. Genital and anal plates with longitudinal sclerotised ridge.
Material examined — 2 females collected 2-III-2012 in mosses, rotting timber and dead leaves on and under fallen tree trunk in tropical rainforest, Saül, Boucle de Monts La Fumée, near confluence of Crique Cochon and Crique quee Hocco, 150 m, 3°37'66''N, 53°12'00''W (sample nr. LM-2012-001), French Guyana. Type preserved in acarological collection of Senckenberg Museum in Goerlitz, paratype (damaged, in parts) in collection of the author.
Description — Measurements – Values of holotype given only, as damaged paratype prevented precise measurements. Total body length 500, prodorsum length 186, prodorsum width between tips of pedotecta I 156, notogaster length 314, notogaster width 186.
Integument – (Figs 3A, 3B, 4C, 4H, 4J). Body robust, colour dark reddish brown. Integument distinctly ornamented, anterior part of prodorsum on prodorsal field (between prodorsal furrow and rostral setae), as well as whole notogaster and ventral plate foveolate, with foveae framed by mostly simple and not undulated margin, some foveolae may have double margins. Size of foveolae variable (mostly 8–15), largest ones (central on notogaster) not larger than half of the length of shortest notogastral setae, their mutual distances equal or smaller than diameter of largest. Some parts of prodorsum (lateral to postbothridial protuberances, axial between setae in and along dorsosejugal furrow smooth, rostral area with fine punctuation. Bothridia with distinct punctuation, narrow strip between bothridia, postbothridial protuberances and tubercles L, as well as epimeral area with dense fine punctuation and cuticle densely, finely penetrated (``chagrenation").
Prodorsum – (Figs 3A, 4A–E). Rostrum conical, broadly rounded, distally with distinct rostral protuberance (rop), this protuberance centrally with lighter, less sclerotised cuticle. Rostral setae (ro) inserted on sides of this protuberance, short (16), curved mediad and ventrad, smooth. Lamellar setae (le) much longer (37), more or less straight, smooth setiform, inserted on short but distinct transversal lamellar ridge. Distal end of setae le almost reaching base of rostral protuberance in dorsal view. Interlamellar setae (in) same or slightly longer in size (43) and similar in shape as setae le, inserted at anterior border of postbothridial protuberances, insertions on small tubercles or areas with strengthened sclerotisation. Bothridium almost rounded, with rather narrow opening. Sensillus smooth, short (36), not longer than setae le and in, distally bifurcate, with short branches attenuated towards the end. Exobothridial seta (ex) shortest of prodorsal setae (10), thin, hardly visible dorsally. Anterior part of bothridial protuberance with distinct transverse anterobothridial ridge, not longer than diameter of bothridium. Prodorsum anterior to this ridge with rather deep prodorsal furrow, causing significant posterior constriction of prodorsal field (area behind setae le). Postbothridial protuberances well developed, almost quadrangular, laterally and posteriorly with distinct postbothridial ridge, posteriorly with single, strong, irregularly spiniform or dentiform tubercle (pbt), reaching anterior margin of notogaster or slightly beyond. Between tubercles, less pronounced and deeper positioned cuticular thickening connects the protuberances. Two distinct semicircular lateral ridges (L) present laterally and slightly posterior to postbothridial protuberances, between postbothridial tubercles and pedotecta II, above dorsosejugal groove,. Lateral outline of prodorsum with slightly pronounced angular pedotectum I, pedotectum II more distinct, triangular or irregular. Blunt projection present also on anterior end of acetabulum II. Acetabulum III dorsally with thickened margins.
Notogaster – (Figs 3A, 4F–H). Regularly oval in general shape, with well pronounced protuberances on posterior end (protuberances bearing setae h3 most developed, followed by those bearing h1 + p1 , p3 and p2 ), but distinct also on lateral sides, diminishing anteriad (largest bearing seta f2 , followed by e2 and d2 ), even setae c2 and c3 on small protuberances. Notogastral setae simple setiform, attenuated distally, relatively short (28–40, e1 longest), setae of rows c, d, e and f not reaching insertions of following more posterior setae. Lateral and posterior setae more curved than others. Some of anterior setae (pair c1 at least) with small, sometimes indistinct basal spur, this often visible only in detailed lateral view (Fig. 4F).
Ventral characters – (Figs 3B, 4I, 4J). Gnathosoma relatively short (86), mentum broader than its length (66 × 46), inverted bell-like, with three blunt projections. Seta a, h, m very short (around 10), smooth, simple. Tectum of podocephalic fossa (tpf) well developed and broad, bluntly rounded, projecting distinctly laterad. Lateral outline of epimeral area with several distinct projections, pedotectum I and II of similar, dentiform or tubercular shape. Posterior to pedotectum I strong, lateral projection covering partly anterior part of acetabulum II, separated from pedotectum I by distinct ridge. Similar, but smaller tubercular projection present also posterior to pedotectum II. Epimeral fields with well visible apodemes, broad and flat, epimeres I largest. Epimeral setal formula 3-1-3-3, epimeral setae simple, setiform and very short (5–10), only 4b and 4c slightly longer (17, 20). Genital plates (72 × 35) with longitudinal ridge, well visible in lateral view, bearing 9 simple setiform genital setae, anterior ones slightly longer (22) than more posterior (14–18). Two pairs of adgenital setae (ag) very short (14–16). Anal plates (86 × 31) with longitudinal sclerotised ridge, bearing 2 pairs of very short (11), simple anal setae, lyrifissure ian not observed. Adanal area well discernible, framed laterally by narrow semicircular ridge, with 3 pairs of adanal setae, these longer than other ventral setae (26–28), simple setiform. Lyrifissure iad present in area of adanal ridges, visible better in lateral view. Lyrifissures ih and ips large, well visible, more-less transverse, positioned anterior to anal opening, mutual distance of ips smaller than distance of ih.
Legs – (Figs 4K, 4L). All legs very strong, robust, with quadrangular or cylindrical segments, genu and tibia of similar length, both about as long as or only slightly longer than broad. Femora I and II proximally with distinct, spur-like projection oriented posteriad. Setae of legs of different shape, those on ventral and lateral side of proximal segments (trochanters, femora, genua and tibiae) setiform, more or less curved, smooth, dorsal setae of the same segments usually stronger, claw-like, curved. Setae of tarsi, particularly at distal end and more on anterior legs than on leg IV, modified, broadened, spiniform or phylliform, with sharp or sometimes bifid end distally, only proral setae usually longer and not broadened, setiform. Trochanteral setal formula 1-1-4-1, setal formula of leg I: 1-5-5[1]-6[1]-24(3), setal formula of leg IV: 1-2-3[1]-5[1]-17. Solenidia small, short (8-10), setiform or bacilliform, slightly curved, only solenidion ω of tarsus I larger, slightly longer (16) than surrounding setae.
Derivatio nominis — The name bifurcata recalls the specific form of bifurcate sensillus, which is very unusual within this genus.
Remarks — Genus Cyrthermannia Balogh, 1958 is known mostly from tropical regions of the world, with some species occuring also in the subtropics. Species of the genus, despite being easily recognized and distinguished, are mostly insufficiently described (with some characters simply missing in descriptions), which makes any analysis of taxonomical relations within the genus quite difficult. Also concepts of the genus have evolved – originally, the genus was proposed for species of Nanhermanniidae, having visible protuberances at least in posterior part of notogaster and with generally short notogastral setae (shorter than distance to the setae positioned in the next posterior row). Later, further genera or subgenera were proposed for species differing in development of the posterior margin of postbothridial ridge. Species with a single strongly developed postbothridial tubercle were left in Cyrthermannia, species with 2 such teeth were placed in the genus Bicyrthermannia Hammer, 1979, and finally species with posterior margin of postbothridial ridge bearing several smaller tubercles similarly as in some species of genus Nanhermannia were placed into Nippohermannia Balogh, 1985, a separate subgenus of Nanhermannia Berlese, 1913. However, separation of genera only on the basis of development of posterior edge of prodorsum may not be sufficient taking into account that Nanhermannia includes both species with a single large tubercle, species with such a tubercle in combination with smaller tubercles, and species with a set of smaller tubercles, and that similarly in Cyrthermannia there are some species which have a few smaller tubercles present laterally to a large tubercle. For further comparison of known species with the new species described herein, I keep the concept of Cyrthermannia as presently defined.
Among the 11 species of Cyrthermannia, as given by Subías (2004, 2018), there are three which I consider either doubtful or not belonging to this genus. Cyrthermannia formosana Tseng, 1982 lacks a single, strongly developed postbothridial tubercle but has a set of numerous small tubercles on the posterior end of postbothridial ridge. Thus, it should be clearly transferred and treated as Nanhermannia (Nippohermannia) formosana (Tseng, 1982) n. comb. Linothrus perinfamis Tseng, 1982 was described on the basis of tritonymph. The figure (Tseng, 1982) indicated that notogastral setae of the nymph are very long, even longer than in some species of Nanhermannia. It is not clear, on what basis Linothrus Tseng, 1982 was synonymised with Cyrthermannia and consequently the species L. perinfamis Tseng, 1982 transferred to this genus by Subías (2004, 2018) as species inquirenda. I consider this placement is doubtful and both genus and species of Tseng need be considered as incertae sedis. As such, it cannot be included in following considerations. Finally, the position of the species Hermannia nasata Warburton, 1912, transferred to Cyrthermannia as species inquirenda by Subías (2004, 2018) remains unclear, as the original description and figure do not provide the necessary information – it is not clear if a strong postbothridial tubercle is or is not present, and prodorsal and all notogastral setae are not mentioned nor depicted. The only indication for placing the species into Cyrthermannia is the overall form of the body with truncate end of notogaster and body surface covered by circular foveolae. As such, the species cannot be included in further analysis and its placement in Cyrthermannia remains questionable.
Thus, disregarding the abovementioned 3 species, 8 species of Cyrthermannia are known. C. bifurcata can be easily distinguished from all of these by the unusual, short and bifurcate sensillus. All other species have a more or less clavate sensillus, with either more distinct, or indistinct and narrow sensillus head. As far as existing information allows analysis, this is not the only difference. First, the new species has setae in inserted on small but distinct tubercles, which were not reported in other species. Second, most if not all species have also at least some epimeral setae distinctly longer, often dilated and sometimes with bristles, while in C. bifurcata, even the longest of epimeral setae are quite short, smooth and setiform.
The new species belongs to the broader group of species with setae le of the same length or only slightly shorter as setae in (for the species included see the determination key below), so that their tips often almost reach insertions of setae ro. Distance of insertions le-ro is therefore usually much less, in some species closer to double length of seta le. Rostral setae are usually positioned on a distinct rostral protuberance or on tubercles. Similarly, several species of this group, including the new species have setae le placed on a short, transverse lamellar ridge. Another, quite remarkable character is indicated for two species of this group and is present on the new species – quite distinct and large, semicircular lateral ridge L, a probable analogue of tubercle L in the lateral postbothridial region, known for example in some Damaeidae (Miko, 2010, 2015). Unfortunately, in many species this area is not depicted with details and there is no mention of this structure – so it is not possible to decide definitely, if this character is specific for the whole group or not.
Within Cyrthermannia, species have 7–9 pairs of genital setae. 9 genital setae appear only within the group mentioned above, however there are also species with 8 genital setae within the group. The only known species with 7 genital setae is C. vicinicornuta Aoki, 1965, which differs also by position of postbothridial tubercles, which are positioned more medially, closer to each other than in other species.
Distribution of other characters seems to be irregular among all known species of the genus, and does not indicate any particular group pattern – the differences are in shape of notogastral setae (smooth setiform vs. broadened to phylliform), presence of small tubercles on lateral part of postbothridial ridge (one to three small tubercles present or absent), and development of cuticular foveolae (round or oval with simple borders vs. modified: irregularly shaped or with doubled or punctuated margins).
Based on analysis of known morphological character, the following determination key distinguishes all known species of Cyrthermannia, including the new species described herein:
Key to the species of the genus Cyrthermannia
1. Setae le much shorter than setae in (less than half of their length), thus le by far not reaching insertions of ro
...... 2
— Setae le of almost the same length as setae in, or only slightly shorter (more than half of their length), le often reaching or almost reaching ro insertions in dorsal view
...... 3
2. Postbothridial ridges lateral to large postbothridial tubercle with 2–3 small tubercles. Notogastral and ventral setae simple, not broadened. Notogastral surface with larger, oval foveolae, their distances smaller than their size. Surface of postbothridial protuberances and epimeres with distinct, dense fine punctuation. Genital plates with 8 setae. Size 540
...... Cyrthermannia baloghorum Pérez-Iňigo and Baggio, 1988
— Postbothridial ridges with single large postbothridial tubercle, without small tubercles. Notogastral and ventral setae broadened. Notogastral surface with medium sized, circular or oval foveolae, their mutual distance usually larger than their diameter. Surface of postbothridial protuberance with few very small foveolae, epimeral area without distinct punctuation. Genital plates with 7 setae. Size 506
...... Cyrthermannia vicinicornuta Aoki, 1956
3. Sensillus short or medium long, clavate, with narrow or more distinct and broadened head, never bifurcate distally. Setae in not inserted on tubercles. At least some epimeral setae long (similar to or slightly shorter than notogastral setae), often broadened and sometimes with short bristles
...... 4
— Sensillus short, distally bifurcate, with two short branches. Setae in inserted on distinct tubercles. Epimeral setae simple, smooth setiform, short or even vestigial, the longest of them significantly shorter than notogastral setae. – Setae ro inserted on distinct rostral protuberance, setae le inserted on transversal short lamellar ridge. Postbothridial area laterally with distinct, semicircular ridge L. Size 500
...... Cyrthermannia bifurcata n. sp.
4. Notogastral setae setiform, not broadened. Notogastral foveolae with distinctly undulated margins
...... 5
— Notogastral setae robust, more or less broadened. Notogastral foveolae simple circular or oval, rarely irregular oval with light dots and interruptions along margins, margins never distinctly undulated
...... 6
5. Setae ro inserted on rostral protuberance. Setae le quite long, with tips clearly reaching behind half of distance to ro insertions, positioned directly on central part of prodorsum. Notogastral foveolae appearing almost stellate, with light dots along margins. Surface of postbothridial protuberances and epimeral area distinctly finely punctuated. Size 568–578
...... Cyrthermannia stellata Balogh, 1970
— Rostral protuberance absent or indistinct. Setae le short, clearly not reaching beyond half of distance to ro insertions, inserted on distinct, short transverse lamellar ridge. Notogastral foveolae resembling shape of flowers. Notogastral setae with distinct spurs. Surface of postbothridial protuberances punctuated, and also with distinct round foveolae. Epimeres without distinct punctuation. Size 478–510
...... Cyrthermannia florens Balogh and Mahunka, 1980
\cledetermination{6. Setae le inserted on transversal short lamellar ridge. Sensillus medium long, longer or as long as seta in, with narrow, not significantly thickened head. Outer margin of circular or oval notogastral foveolae smooth, without interruptions or luminous dots} {7}
— Setae le inserted directly on prodorsal surface, lamellar ridge absent. Sensillus short, shorter than length of setae in, with short peduncle and distinct, broadened head. Notogastral foveolae irregularly oval, with outer margins interrupted and/or with luminous dots. Notogastral and prodorsal setae blade-like, sharp, with distinct spurs. Size 500
...... Cyrthermannia luminosa Hammer, 1971
7. Notogastral setae simple blade-like, mostly with spurs, short – seta c1 not reaching insertion of d1 . Setae ro inserted directly on rostrum, or on individual tubercles, rostral protuberance absent. Notogastral foveolae round or slightly irregular oval, with simple smooth margins. Notogaster without humeral tubercles
...... 8
— Notogastral setae broadened, with a velum on both sides, long - seta c1 reaching distinctly posterior of insertion of d1 . Setae ro inserted on well developed rostral protuberance. Notogastral foveolae round, with smooth external margin and with irregular internal line. Notogaster with distinct humeral tubercles. Size 487–535
...... Cyrthermannia tuberculata Balogh, 1958
8. Setae ro inserted on distinct individual tubercles. Notogastral and prodorsal setae only slightly broadened. Sensillus with relatively broad head. Genital plates with 8 setae. Adanal setae setiform. Size 508–525
...... Cyrthermannia simplex Mahunka, 1985
— Setae ro inserted on rostral surface, not on tubercles. Notogastral and prodorsal setae strongly broadened. Sensillus with narrow head, not significantly broader than stalk. Genital plates with 9 setae. Adanal setae long and distinctly broadened, phylliform. Size 502–535
...... Cyrthermannia guadeloupensis Mahunka, 1985
References
Badejo, M. A., Woas, S., Beck, L. 2002. Redescription of Archegozetes magnus (Sellnick, 1925) (Trhypochthonioidea) from Brazil and description of two new species of nanhermanniid mites: Bicyrthermannia nigeriana and Masthermannia seropedica (Nanhermannioidea) (Acari: Oribatida). Genus, 14 (1): 125-149.
Balogh, J. 1958. Oribatides nouvelles de l'Afrique tropicale. Rev. zool. bot. afr., LVIII, 1-2: 1-34.
Balogh, J. 1972. The Oribatid genera of the world. Akadémiai Kiadó Budapest, 188 pp.
Balogh, J., Balogh, P. 1992. The Oribatid mites genera of the world. Volume I. Hungarian National Museum Press, 263 pp.
Balogh, P. 1997 New species of oribatids (Acari) from the Neotropical Region. Opusc. Zool. (Budapest), 29/30: 21-30.
Colloff, M.J. 2013. Species-groups and biogeography of the oribatid mite family Malaconothridae (Oribatida: Malaconothroidea), with new species from the south-western Pacific region. Zootaxa, 3722 (4): 401-438. doi:10.11646/zootaxa.3722.4.1 ![]()
Colloff, M.J., Cameron, S.L. 2013. A phylogenetic analysis and taxonomic revision of the oribatid mite family Malaconothridae (Acari: Oribatida), with new species of Tyrphonothrus and Malaconothrus from Australia. Zootaxa, 3681: 301-346. doi:10.11646/zootaxa.3681.4.1 ![]()
Ermilov, S. G., Starý, J. 2019. New taxa of oribatid mites (Acari, Oribatida) from the Korup national park, Cameroon. The genus Malaconothrus Berlese 1904 (Malaconothridae). Zool. Zh., 98 (5): 504-512. doi:10.1134/S004451341903005X ![]()
Hammer, M. 1979. Investigations on the Oribatid fauna of Java. Det Kongelige Danske Videnskabernes Selskab. Biol. Skrift., 22, 9: 1-126.
Mahunka, S. 1992. New and interesting mites from the Geneva Museum LXIII. A survey of the oribatid fauna of Senegal (Acari, Oribatida). Rev. Suisse Zool., 99: 673-712. doi:10.5962/bhl.part.79846 ![]()
Miko, L. 2010. Taxonomy of European Damaeidae (Acari: Oribatida) III. Species of the Kunstidamaeus tenuipes (Michael, 1885) group, with a description of Kunstidamaeus fraterculus n. sp. from East Slovakia. Zootaxa 2327: 51-64. doi:10.11646/zootaxa.2327.1.4 ![]()
Miko, L. 2015. Taxonomy of European Damaeidae VIII. Contribution to classification of genus Damaeus C. L. Koch, 1835, with a review of Adamaeus Norton, 1978 and Paradamaeus Bulanova-Zachvatkina, 1957 and redescription of three species. Zootaxa 3980 (2): 151-188. doi:10.11646/zootaxa.3980.2.1 ![]()
Miko, L. 2016. Oribatid mites (Acarina, Oribatida) from French Guyana: review of the genus Rhynchoribates and description of three new species. Zootaxa, 4061 (2): 131-145. doi:10.11646/zootaxa.4061.2.3 ![]()
Norton, R., Behan-Pelletier, V. 2009. Oribatida. In: Krantz, G.W., Walter, D.E. (Eds.), A manual of acarology. 3rd Edition. Edited by. Texas Tech University Press, Lubbock, Texas, 430-564.
Subías, L.S. 2004. Listado sistemático sinonímico y biogeográfico de los ácaros oribátidos (Acariformes, Oribatida) del mundo (1758-2002). Graellsia, 60 (supplement): 3-305. Updated February, 2011. Available from: http://www.ucm.es/info/zoo/Artropodos/Catalogo.pdf ![]() (Accessed 9 July 2011) doi:10.3989/graellsia.2004.v60.iExtra.218
(Accessed 9 July 2011) doi:10.3989/graellsia.2004.v60.iExtra.218 ![]()
Subías, L.S. 2018. Listado sistemático sinonímico y biogeográfico de los ácaros oribátidos (Acariformes, Oribatida) del mundo (1758-2002). Updated on May 2013. Available from: www.ucm.es/info/zoo/Artropodos/Catalogo.pdf ![]() (accessed September - December 2018)
(accessed September - December 2018)
Tseng, Y-H. 1982. Taxonomical study of oribatid mites from Taiwan (Acarina: Astigmata) (I). Chin. J. Ent., 2 (1): 53-106.
Weigmann, G. 1997. Systematics of Central European families and genera of Malaconothroidea (Acari, Oribatida). Abh. Ber. Nat. Görlitz 69 (6): 3-10.
Weigmann, G. 2006. Hornmilben (Oribatida). Die Tierwelt Deutschlands, 76. Teil. Goecke and Evers, Keltern, 520 pp.
Yamamoto, Y. 1996. Two new oribatid mites of the family Malaconothridae in Wakayama Prefecture, Central Japan (Acari: Oribatei). Acta Arachnol., 45: 119-124. doi:10.2476/asjaa.45.119 ![]()
Yamamoto, Y., Aoki, J.-I. 1997. A new oribatid mites of the genus Malaconothrus from Irimote Island, Southwest Japan (Acari: Oribatei: Malaconothridae). J. Acar. Soc. Jap., 6: 43-47. doi:10.2300/acari.6.43 ![]()



2019-05-09
Date accepted:
2019-08-06
Date published:
2019-08-09
Edited by:
Baumann, Julia

This work is licensed under a Creative Commons Attribution 4.0 International License
2019 Miko, Ladislav
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)