




The first DNA barcodes for the Australian platypus tick Ixodes ornithorhynchi Lucas, 1846 (Acari: Ixodidae) to facilitate conservation efforts for a declining parasite and its host
Kwak, Mackenzie L.1 ; Griffiths, Josh2 ; Barry, Dylan3 ; Begent, Mikayla4 ; Hoang, Tyrone5 ; Taafua, Luisa6 and Chiovitti, Anthony7
1✉Evolutionary Biology Laboratory, Department of Biological Science, National University of Singapore, 16 Science Drive 4, 117558, Singapore.
2CESAR, 293 Royal Parade, Parkville Victoria 3052 Australia.
3Gene Technology Access Centre (GTAC), 77 Story Street, Parkville, Victoria 3052 Australia.
4Gene Technology Access Centre (GTAC), 77 Story Street, Parkville, Victoria 3052 Australia.
5Gene Technology Access Centre (GTAC), 77 Story Street, Parkville, Victoria 3052 Australia.
6Gene Technology Access Centre (GTAC), 77 Story Street, Parkville, Victoria 3052 Australia.
7Gene Technology Access Centre (GTAC), 77 Story Street, Parkville, Victoria 3052 Australia.
2018 - Volume: 58 Issue: 4 pages: 845-849
https://doi.org/10.24349/acarologia/20184287ZooBank LSID: 65AEFA11-8A3F-425B-B016-A7FBBA8BBCF9
Keywords
Abstract
Ixodes ornithorhynchi is a host-specific tick of the Australian platypus (Ornithorhynchus anatinus) (Roberts, 1970). In recent decades, many platypus populations have become fragmented and both the host and parasite are believed to have gone extinct on the mainland in at least one Australian state (Grant and Temple-Smith, 2003). The platypus has an extensive range spanning most of eastern Australia, but both it and I. ornithorhynchi are of conservation interest due to this pattern of decline (Menkhorst and Knight, 2011). Although tick population sizes can be monitored using morphological identification, assessing the conservation status of a tick population often requires molecular study to identify levels of genetic diversity and instances where genetic bottlenecks form. To better facilitate research on the conservation biology of I. ornithorhynchi and the platypus in Australia, the first DNA barcodes of I. ornithorhynchi are presented.
Twelve tick specimens (representing 9 nymphs and 3 adult females) collected from two platypodes (O. anatinus) in Jackson Creek, Sunbury, Victoria, Australia were identified using morphology-based keys provided by Roberts (1970) under a light microscope. Scanning electron micrographs were taken of specimens from which DNA barcodes were generated. These were taken using a Hitachi TM3030 electron microscope and are presented in Figure 1.


Total genomic DNA was extracted from 12 individuals using the Nucleospin Tissue Kit (Machery-Nagel, Germany) following the manufacturer’s protocols. The mitochondrial COI gene was targeted with a primer combination adapted from Kwak et al. (2017). This comprised a forward primer, HCO2064 (5’-GGT GGG CTC ATA CAA TAA ATC C-3’) and a reverse primer, HCO1240 (5’-CCA CAA ATC ATA AAG ACA TTG G-3’) yielding a fragment of 846 bp in length. The COI gene segment was amplified from the purified genomic DNA using the Phusion HotStart Taq DNA polymerase (New England Biolabs, USA) with accompanying reagents according to the manufacturer’s protocols. The PCR was run on a BioRad model T100 thermocycler with a temperature cycle encompassing denaturation at 98 °C for 30 s, annealing at 48 °C for 30 s, and primer extension at 72 °C for 45 s. The cycle was repeated 30 times and bracketed by an initial denaturation step at 98 °C for 2 min and a terminal extension step at 72 °C for 5 min. PCR products were visualised by gel electrophoresis (1%), stained using SYBRSafe (Invitrogen, USA) and compared against size standards (Quick Load PCR Marker N0475S, New England Biolabs, USA). PCR products were purified using the QIAquick PCR Purification Kit (Qiagen, Germany) following the manufacturer’s protocol and were submitted to the Australian Genome Research Facility (AGRF) for bidirectional Sanger sequencing using the same primers as those used for PCR.
A phylogenetic tree was constructed using the program MEGA6 (Tamura et al., 2013). Sequences were trimmed then aligned using the ClustalW function. The phylogeny was inferred using the neighbour-joining method with 10,000 bootstrap replications. Sequences of I. ornithorhynchi were compared against those of other Ixodes species present on GenBank and are presented in Table 1. Species used for comparison include those from the main Australian subgenera as well as the Afrixodes and Partipalpiger which exhibit morphological similarities with I. ornithorhynchi, as listed by Clifford et al. (1973). The southern reptile tick (Bothriocroton hydrosauri) was used as an outgroup (GenBank reference: FJ584427.1). Bootstrap values <45 are not shown as the purpose of the phylogeny is to demonstrate the distinctiveness of the species from its relatives rather than to infer, and support, deeper evolutionary relationships.
The adult female specimens were identified as I. ornithorhynchi based on a combination of morphological features including unarmed coxae, the absence of cornua and auriculae, large porose areas, hypostomal dentition of 3/3, long palps in which all four articles are distinct and without spurs, and an anal groove which does not meet posteriorly. Nymphs were identified as I. ornithorhynchi based on a combination of morphological features including unarmed coxae, the absence of cornua and auriculae, hypostomal dentition of 2/2, long palps in which all four articles are distinct and without spurs, and an anal groove which does not meet posteriorly.
Unique sequences were obtained at the COI locus for two I. ornithorhynchi specimens representing one nymph and one adult female tick (Table 1). All other sequences were identical to that of the nymph.


Phylogenetic analysis (Figure 2) placed I. ornithorhynchi in a distinctly separate clade from all other tick species to which it was compared. Although several morphological similarities exist between I. ornithorhynchi and members of other subgenera, I. ornithorhynchi does not fall within any of these clades.

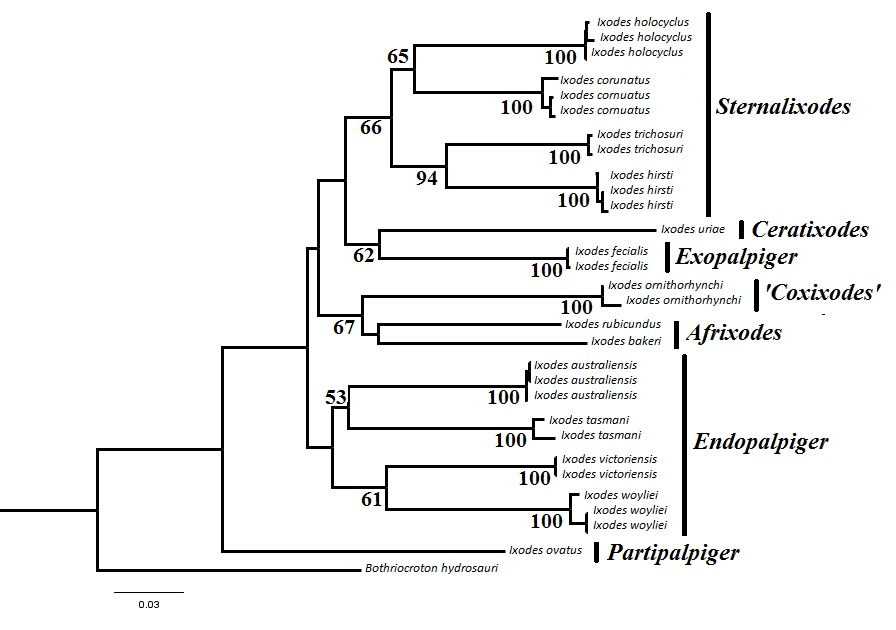
Based on the phylogenetic tree presented in Figure 2, I. ornithorhynchi appears to be distinct from all other species to which it was compared. This is unsurprising, as the higher classification of I. ornithorhynchi has been far from settled historically due to the morphological distinctiveness of the species. Since Schulze (1941) erected the monotypic genus Coxixodes for I. ornithorhynchi in a short passage of German, few taxonomists have accepted and used it, even though it was reclassified as a subgenus. Roberts (1970) completely ignored the subgenus and Clifford et al. (1973) avoided the use of Coxixodes, citing limited available data to support its validity.
The evolutionary relationship between I. ornithorhynchi and the other Ixodes subgenera remains poorly understood. The genus Ixodes comprises over 240 known species, many of which are still without molecular data (Guglielmone et al., 2010). This lack of data renders a total comparison of species at the COI locus impossible. As more DNA sequences of the COI locus become available for members of Ixodes, it may become possible to determine the evolutionary relationships between closely related Ixodes species. However, as demonstrated by Kwak et al. (2017), the COI marker only appears useful in distinguishing the evolutionary relationships between closely related members of the Ixodes genus. Therefore, the phylogeny presented in Figure 2 is only valuable to demonstrate the distinctiveness between I. ornithorhynchi and the other Ixodes to which it was compared. Analysis of more highly conserved genetic markers will be required to make inferences about the evolutionary relationships amongst species of this highly diverse genus.
Although DNA barcodes of I. ornithorhynchi are useful for delimiting species boundaries, they may also serve a purpose in future attempts to monitor and map the population dynamics of platypus ticks and their hosts, both past and present. Nieberding and Olivieri (2007) suggested that parasites could be used to map past population events in their hosts such as rapid range expansions or contraction events causing genetic bottlenecks. Biek et al. (2006) successfully demonstrated this when they showed how an examination of a directly transmitted parasite could be used to map the recent demographic history of its host population. As I. ornithorhynchi is a directly transmitted, host specific parasite, and the only tick species regularly found on the platypus, it is a good candidate for such study. Haplotype diversity at the COI locus could be used as a proxy to estimate historical levels of gene flow between platypus populations. Although CO1 is a maternaly inherited marker, it may still prove useful for providing a raw estimation of population stability. Previously, COI has been used in New Zealand in endangered Amblyomma sphenodonti populations to estimate gene flow and intrapopulation genetic diversity (Miller et al., 2007). Measuring haplotype diversity at the COI locus could similarly be a valuable way to infer and monitor gene flow and relative genetic diversity in I. ornithorhynchi populations to ensure the continued survival of one of Australia’s most cryptic arthropods across its range.
Biek R., Drummond AJ. and Poss M. 2006. A virus reveals population structure and recent demographic history of its carnivore host. Science, 311(5760): 538-541. doi:10.1126/science.1121360 ![]()
Clifford CM., Sonenshine DE., Keirans JE. and Kohls GM. 1973. Systematics of the subfamily Ixodinae (Acarina: Ixodidae). 1. the subgenera of Ixodes. Annals of the Entomological Society of America 66(3): 489-500. doi:10.1093/aesa/66.3.489 ![]()
Grant TR. and Temple-Smith PD. 2003. Conservation of the platypus, Ornithorhynchus anatinus: threats and challenges. Aquatic Ecosystem Health & Management 6(1): 5-18. doi:10.1080/14634980301481 ![]()
Guglielmone AA., Robbins RG., Apanaskevich DA., Petney TN., Estrada-Peña A., Shao R. and Barker SC. 2010. The Argasidae, Ixodidae and Nuttalliellidae (Acari: Ixodida) of the world: a list of valid species names. Zootaxa 2528: 1-28.
Kwak ML., Beveridge I., Koehler AV., Malipatil M., Gasser RB. and Jabbar A. 2017. Phylogenetic analysis of the Australasian paralysis ticks and their relatives (Ixodidae: Ixodes, Sternalixodes). Parasites & Vectors 10(1): 122. doi:10.1186/s13071-017-2045-4 ![]()
Menkhorst P. and Knight F. 2011. Field guide to the mammals of Australia (3rd Ed). Melbourne: Oxford University Press.
Miller HC., Conrad AM., Barker SC. and Daugherty CH. 2007. Distribution and phylogenetic analyses of an endangered tick, Amblyomma sphenodonti. New Zealand Journal of Zoology 34(2): 97-105. doi:10.1080/03014220709510068 ![]()
Nieberding CM. and Olivieri I. 2007. Parasites: proxies for host genealogy and ecology?. Trends in Ecology & Evolution 22(3): 156-165. doi:10.1016/j.tree.2006.11.012 ![]()
Roberts FHS. 1970. Australian ticks. Melbourne: CSIRO Publishing.
Schulze P. 1941. Das Geruchsorgan der Zecken. Untersuchungen über die Abwandlungen eines Sinnesorgans und seine Stammesgeschichtliche Bedeutung. Zeitschrift für Morphologie und Ökologie der Tiere 37: 491–564.
Tamura K., Stecher G., Peterson D., Filipski A. and Kumar S. 2013. MEGA6: molecular evolutionary genetics analysis version 6.0.. Molecular biology and evolution 30(12): 2725-2729. doi:10.1093/molbev/mst197 ![]()



2017-12-21
Date accepted:
2018-08-08
Date published:
2018-09-21
Edited by:
McCoy, Karen

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Kwak, Mackenzie L.; Griffiths, Josh; Barry, Dylan; Begent, Mikayla; Hoang, Tyrone; Taafua, Luisa and Chiovitti, Anthony
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)