




Oribatid mites of conventional and organic vineyards in the Valencian Community, Spain
Seniczak, Anna1 ; Seniczak, Stanisław2 ; García-Parra, Ivan3 ; Ferragut, Francisco4 ; Xamaní, Pilar5 ; Graczyk, Radomir6 ; Messeguer, Enric7 ; Laborda, Rafael8 and Rodrigo, Eugenia9
1✉ Department of Biology and Animal Environment, UTP University of Science and Technology, ul. Hetmańska 33, 85-039 Bydgoszcz, Poland.
2Department of Evolutionary Biology, Faculty of Natural Sciences, Kazimierz Wielki University, Ossolińskich 12, 85-092 Bydgoszcz, Poland.
3Dpto. de Ecosistemas Agroforestales, Universitat Politècnica de València, Camino de Vera s/n, 46022 València, Spain.
4Instituto Agroforestal Mediterráneo, Universitat Politècnica de València, Camino de Vera s/n, 46022 València, Spain.
5Departamento de Ecosistemas Agroforestales, Universitat Politècnica de València, Camino de Vera s/n, 46022 València, Spain.
6Department of Biology and Animal Environment, UTP University of Science and Technology, ul. Hetmańska 33, 85-039 Bydgoszcz, Poland.
7Dpto. de Ecosistemas Agroforestales, Universitat Politècnica de València, Camino de Vera s/n, 46022 València, Spain.
8Instituto Agroforestal Mediterráneo, Universitat Politècnica de València, Camino de Vera s/n, 46022 València, Spain.
2018 - Volume: 58 Issue: Suppl pages: 119-133
https://doi.org/10.24349/acarologia/20184281Keywords
Abstract
Spain is the country with the largest area of vineyards in the world (with more than 1 million ha, i.e. 14% of world’s total vineyard area) (OIV 2016). It also leads in the organic viticulture, with the largest area of organic vineyards (MAGRAMA 2015). The main organic wine producing regions in Spain are Castilla La Mancha (4,095 ha), Murcia (3,722 ha), València (2,086 ha) and Cataluña (1,127 ha) (Fabeiro et al. 2007).
The main difference between conventional and organic (also called ecological) agriculture is the use of pesticides and chemical fertilizers in conventional farming and prohibition of these activities in organic systems. The organic production relies on crop rotation, fixation of natural nitrogen, biologically active soil, recycled farm manure and crop residues, and on biological or mechanical weed and pest control (Mäder et al. 2002; Bengtsson et al. 2005). In Spain in organic vineyards some insecticides and fungicides are allowed (azadirachtin, sulfur, and sometimes, when the weather conditions are optimal for some fungi, copper calcium sulfate), while in conventional vineyards different pesticides are used (including chlorpyrifos, azoxistrobin, mancozeb).
The biological activity of soil is affected by oribatid mites (Walter and Proctor 1999), which are one of the most abundant arthropod group in the organic horizons, also in vineyards (Suzuki 1979), where they play an important role in decomposition of organic matter, nutrient cycling and soil formation (Behan-Pelletier 1999). Additionally, some species can possibly serve in the pest control. For example, the pathogenic fungus, Rhizoctonia solani that attacks grapevine roots (Walker 1992), can be controlled by the oribatid species Protoribates agricola and Scheloribates azumaenis (Nakamura et al. 1991; Enami and Nakamura 1996).
In undisturbed agroecosystems the density of Oribatida easily reaches several thousand individuals per 1 m2 and 20-50 species (Behan-Pelletier 1999). These mites are more abundant in grasslands, with the densities up to several hundred thousand individuals per 1 m2. In the agricultural fields the density of Oribatida is about ten times lower, mainly because of the cultivation practices that reduce the density of the soil fauna (Niedbała 1980). Among these practices, especially mechanical works on the soil, like tillage, ploughing, disking, etc. or using too high amounts of fertilizers or some chemicals, can have negative effects on the oribatid communities (summarized by Behan-Pelletier 1999).
The Oribatida of vineyards are poorly studied in comparison to other agroecosystems, like meadows, pastures, orchards or arable fields. Some studies on the mites from vineyards have been conducted in Germany (Jörger 1991) and Brazil (Johann et al. 2014), but the Oribatida were treated only as a group. More detailed studies on these mites from vineyards have been carried out in Japan (Suzuki 1979), Azerbaijan (Samedov et al. 1987), Italy (Nannelli and Simoni 2002) and India (Acharya and Basu 2014), but they were based only on adults, and none of them included organic vineyards.
Most studies showed a positive effect of organic farming on density (96 of 117 studies; i.e. 82%) and species richness (53 of 63 studies; i.e. 84%) of plants and animals (Bengtsson et al. 2005). Regarding microarthropods, most studies on the effect of organic and conventional agriculture have been carried out in the annual cultures (e.g. Bettiol et al. 2002; Van Leeuwen et al. 2015) and more soil microarthropods were found in organically managed farming systems than in conventional ones. Oribatida reacted positively to organic management, together with Uropodina (Badejo et al. 2004). However, in apple orchards that were permanent crop, there was no significant difference in density of oribatid mites between the organic and conventional management (Doles et al. 2001). Based on these results we hypothesized that organic cultivation of vineyards vs. conventional one has no impact on the oribatid density and species richness.
The aims of this study were (1) to compare the oribatid communities in conventional and organic vineyards, (2) investigate the community structure of Oribatida in selected habitats in vineyards, (3) compare the dynamics of these mites in autumn and spring, and (4) improve the knowledge of the species diversity in the vineyards.
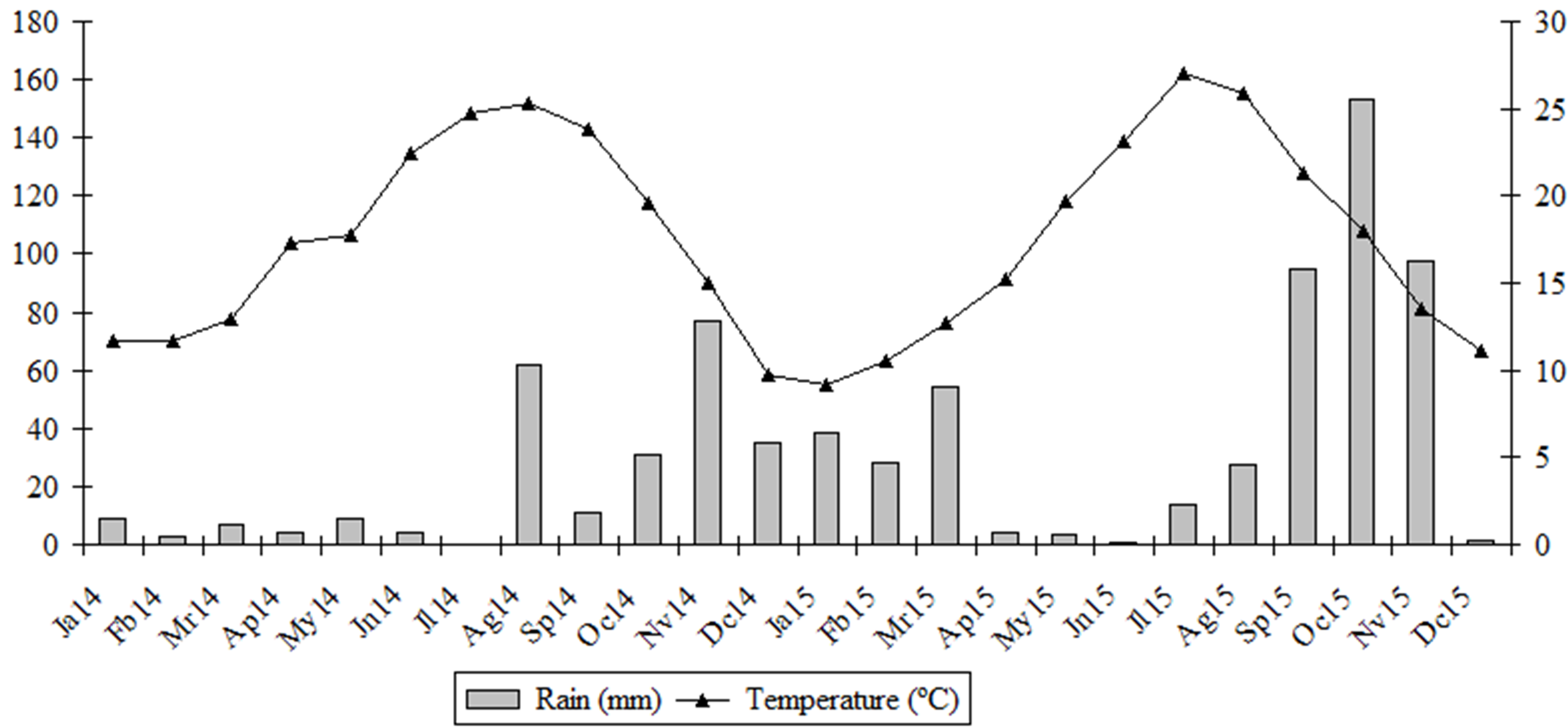
The study was carried out in El Poble Nou de Benitatxell, a village located in the Valèncian Community, Spain (Figure 1). The village has an area of 12.65 km2 and is situated on a hill of the elevation 159 m a.s.l. The climate of the area is clearly Mediterranean, with mild temperature in winter and higher in summer, with a characteristic period of drought in summer and higher rainfall in spring and autumn (Figure 2). The year 2014 was drier (annual rainfall 251 mm) than the year 2015 (518 mm). In the sampling seasons (autumn 2014 and spring 2015), the average temperature was 17°C and 18°C, respectively, and the average rainfall was 135 mm and 25 mm (MAGRAMA 2015).



The soils are calcareous, including deposition and lithology marls (‘Tap” facies) and decarbonated soils (terra rossa) (Barbér Vallés and Moity Martín 2009). The morphology of the landscape is characterized by a strong anthropization, predominantly terraced with stone walls. Traditionally, these terraces were planted with cereals, legumes (beans) and Muscat grapes, but nowadays unirrigated vineyards with table grapes (Muscat of Alexandria variety) predominate (80% of the cultivated land, i.e. 368 ha; Statistics National Institute: Agrarian Census 2009).
Most vineyards have been cultivated in a conventional way, with the use of synthetic fertilizers and pesticides. But since farmers are becoming aware of problems caused by these substances in the ecosystems, they have started an initiative to change the conventional system into organic one. Thus, the crop receives water exclusively from the rain and the farmers maintain the traditional cultural techniques without mineral fertilizing and only some pesticides (azadirachtin, sulfur, and seldom, copper calcium sulfate). In Spain the normative says a farmer needs 4 years to get the organic national certification. The project also prioritizes fair trade, giving a fair price to the farmer and selling locally, thus reducing CO2 emissions form the transport (Domingo et al. 2014).
Four study sites, which are included in this local project of changing the table grapes production into organic system, were selected. They were treated as replicates and each included a conventional vineyard, an organic vineyard and a natural habitat (control), that was in plots 1-3 an abandoned vineyard and in plot 4 the area never used in agriculture. Their locality was respectively: site 1 ˗ 38°44'16.28" N, 0º7'33.80" E; 38º44'16.20" N, 0º7'32.35" E; 38º44'17.52" N, 0º7'33.35" E, site 2 ˗ 38º44'15.59" N, 0º8'22.83" E; 38º44'13.17" N, 0º8'24.57" E, and 38º44'11.74" N, 0º8'24.97" E), site 3 ˗ 38º43'38.10" N, 0º8'51.19" E; 38º43'38.34" N, 0º8'50.50" E; 38º43'40.63" N, 0º8'54.48" E, and site 4 ˗ 38º43'5.94" N, 0º8'17.71" E; 38º43'50.83" N, 0º7'48.42" E, 38º43'52.24" N, 0º7'48.48" E. The vines in sites 1-4 were planted respectively in the years 1992, 1967, 1980 and 1987, but since 2012 all of them have been cultivated in organic system.
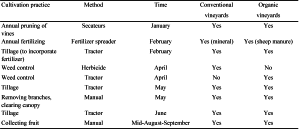
In the conventional system of management (Table 1) the farmers apply herbicides to kill weeds before spring (the active ingredient is glyphosate 36%) and they use chemical fertilizers with different formulations. In organic vineyards they do not apply any herbicide, but from the second tilling on, an implement that cuts weeds between vines is added. Since 2013 these plots are fertilized with sheep manure. Finally, uncultivated plots do not show any intervention or modification, being occupied by spontaneous vegetation.
In total, 40 plant species were recorded in all plots during a 6-month study (March-August). The number of species was similar in conventional and organic vineyards and in the natural habitat (Table 2), while the plants density and plant cover were highest in the natural habitat, followed by the organic vineyards, and were lowest in conventional vineyards.


Sampling was carried out in autumn (13 Nov. 2014) and spring (12 Jun. 2015). In each of the four sites two parallel samples (each of the area 950 cm2, 10 cm deep) were collected with metal corer from seven plots: (1) natural habitat, treated as the control, (2) conventional vineyard, zone between vine rows, driven by a tractor (hereafter referred to as Tr), (3) conventional vineyard, zone between vines (Vi), (4) border of conventional vineyard (Bo), and organic vineyard from the same habitats, (5) Tr, (6) Vi and (7) Bo. In total 112 samples were collected (4 sites x 2 samples x 7 plots x 2 seasons).
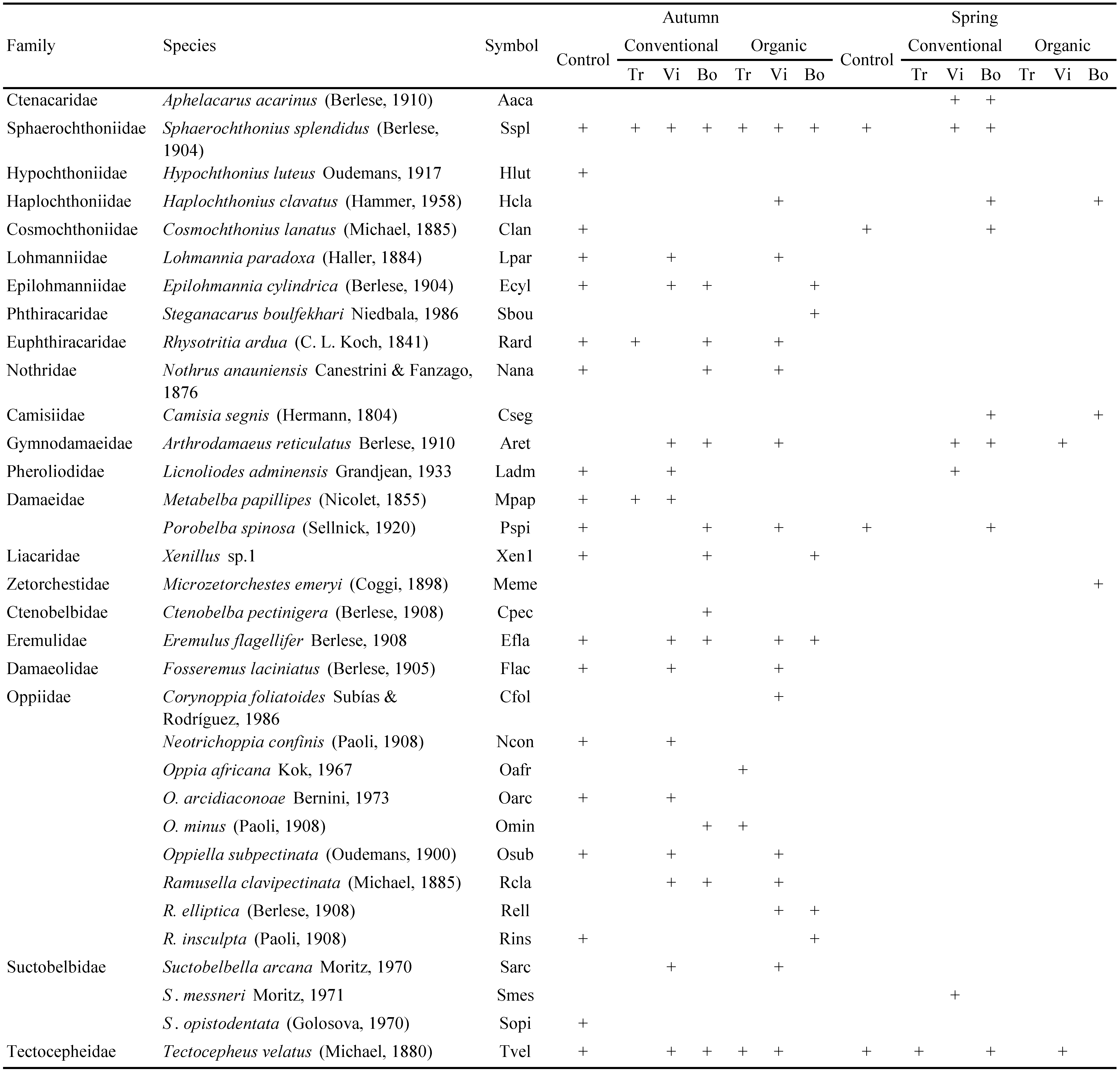
The samples were extracted in Berlese funnels during 4 days at the Universitat Politècnica de València; the time of extraction was based on the observation that after 4 days no living mites were present in the samples. The mites were preserved in 70% ethanol and Oribatida were sorted out under stereomicroscope and determined, including the juveniles, at the UTP University of Science and Technology. The nomenclature of the Oribatida follows Weigmann (2006) and partly Subías (2004, 2015). The data presented in tables 3 and 5 and in Figure 3 refer to all mites, including adults and juveniles. Authors and dates of the taxonomic names are listed in Table 1.
Oribatid mite populations were characterized by the density (A), dominance (D) and constancy (C) indices, and oribatid mite communities were characterized by the number of species (S) and the Shannon (H’) diversity index (Odum 1982). The basic statistical descriptors included the minimum, maximum, mean values and standard deviation. For the other statistical analyses, the values were log-transformed ln (x+1) (Łomnicki 2010). Normality of the distribution was tested with the Kolmogorov-Smirnov test, while the equality of variance in different samples, with the Levene test. The assumption of normality or equality of variance was not met, so the non-parametric ANOVA rang Kruskal-Wallis was used and then, in case of significant differences between averages, the multiple comparison test between average ranks was used. The level of significance for all statistical tests was accepted at α = 0.05. The statistical calculations mentioned above were carried out with STATISTICA 10.0 software.
Subsequently, based on the same log-transformed ln (x+1) data set the analysis of the community structure of the Oribatida with detrended correspondence analysis (DCA) was performed using MVSP 3.2 (Multi Variate Statistical Package, Kovach Computing Services; Piernik 2008). DCA, instead of PCA, was carried out because the length of gradient was 3.9 indicating that unimodal models are more appropriate than linear models (Leps and Smilauer 2003), because the structure of the data has unimodal character (i.e. each species occurs in particular range of a given habitat parameter) (Hill and Gauch 1980).
In total 3,225 oribatid mites were obtained, represented by 59 species from 39 families (Table 3). Oppiidae were represented by nine species, Oribatulidae by eight species, Suctobelbidae by three species, while other families were represented by one or two species.



The average density of Oribatida in the control was over 2-fold higher (5,900 individuals per 1 m2) than in the vineyards (average value from all habitats was 2,820 individuals per 1 m2 in conventional vineyards and 2,250 individuals per 1 m2 in organic vineyards), but these results were insignificant (Table 4). In each type of vineyard the significant differences were only observed between plots Tr and Vi in the autumn (Table 4).


In the control, the density of Oribatida was similar in both seasons, while in the vineyards, both conventional and organic, it was several-fold higher in autumn than in spring (Table 4). The decrease of abundance was especially conspicuous in the juveniles (Table 4); in the autumn their participation among Oribatida was on average 11% and in spring 6%. In all plots the number of species was higher in autumn than in spring, and the Shannon diversity index usually followed this pattern.
According to detrended correspondence analysis (DCA), oribatid mite communities of the control were clearly different from those of the vineyards (Figure 3). The conventional and organic vineyards did not differ from each other, but the three studied habitats differed. Among more abundant species, Oribatula excavata mostly structured the ordination and was characteristic for vineyards (Figure 3). Another species that also affected the ordination was Galumna tarsipennata that was present in conventional and organic vineyards, as well as in the control, but its abundance was significantly higher in zone Vi than in other habitats (Table 5).


In the vineyards, Oribatula excavata dominated, followed by Minunthozetes quadriareatus and Passalozetes africanus, while in the control these species were not abundant (Table 5). In the control the most abundant was Oppiella subpectinata followed by Eremulus flagellifer which were by contrast not abundant in the vineyards. Based on the list of Subías and Shtanchaeva (2012), Podoribates longipes and Steganacarus boulfekhari are new to the Spanish fauna.




The density and species diversity of Oribatida was similar in both types of vineyards, despite the fact that the vegetation was more abundant and diverse in the organic system. The herbicides used in conventional vineyards clearly affected the vegetation there, but not the soil Oribatida. Generally herbicides and fungicides are less harmful to oribatid mites than are insecticides (Lebrun 1977). When they are applied occasionally and in doses permitted in the agriculture, they do not change significantly the density of Oribatida (Niedbała 1980; Fuangarworn et al. 2002). In contrast, insecticides decrease the general density of Oribatida, but some species tolerate them, including Oppiella nova, Scheloribates latipes and S. laevigatus. The density of these species increases (Niedbała 1980; Adán et al. 1991), partly due to lower abundance of their predators, caused by insecticides (Menhinick 1962). In contrast, Galumna tarsipennata reacted negatively to pesticides (Adán et al. 1991), but in our study it had a similar abundance in both types of vineyards, that means it is tolerant of herbicides used in conventional vineyards.
This reaction of oribatid mites contrasts with the general findings that organic farming has a positive effect on the abundance and species richness of organisms (summarized by Bengtsson et al. 2005), but is consistent with observations on the oribatid mites in apple orchards (Doles et al. 2001) and on other arthropods in vineyards (Meseguer Cervera 2014; García-Parra 2015). In the same vineyards of El Poble Nou de Benitatxell as studied here, the abundance of thrips (Thysanoptera), eulophids (Eulophidae), aphelinids (Aphelinidae) and aphidiins (Aphidiinae) did not differ between both types of cultivation (Meseguer Cervera 2014), while the abundance of mesostigmatic mites was higher in conventional vineyards than in organic ones (García-Parra 2015).
In both types of vineyards the density of Oribatida and their species richness varied between three studied habitats (zone between vine rows, zone between vines, border of vineyard), which can be mainly explained by the sensitivity of these mites to the mechanical treatment of soil (Behan-Pelletier 1999). For example tillage reduced the density of Oribatida, but did not affect Mesostigmata, and increased the density of Prostigmata (Winter et al. 1990). Also in the vineyards of El Poble Nou de Benitatxell, the cultural practices did not affect the density of Mesostigmata, but their species diversity (García-Parra 2015). These differences between Oribatida and Mesostigmata can be explained by their different mobility; the predatory Mesostigmata move quickly and easily colonize new habitats, while Oribatida are slowly moving, 11-20 cm a day (Berthet and Gerard 1965), being more sensitive than the Mesostigmata to changes in their environment.
The density of Oribatida in the present study was similar to that reported from the vineyards in Italy (3,300-6,900 adult individuals per 1 m2; Nannelli and Simoni 2002), and seems to be typical for these ecosystems in the Mediterranean region. In other geographical regions, much higher densities of Oribatida were noted in vineyards; to a great extent this can be explained by different climatic conditions, especially higher humidity or colder climate. For example, in Japan, where grapes grow in monsoonal climate, with the annual rainfall above 1000 mm, including 800 mm in growing season, the density of Oribatida in vineyards was 57,000 individuals per 1 m2 (Suzuki 1979). In Azerbaijan the density of Oribatida varied depending on the soil type between 40,000-422,000 individuals per 1 m2 (Samedov et al. 1987). Although the wine regions of Azerbaijan are characterized by a low rainfall (250-600 mm), similarly to those in Spain, in many vineyards irrigation is used, and the average annual temperatures are lower than in Spain (10.5-15.5°C), that seems to provide more favorable conditions for oribatid mites.
In the control the density of Oribatida was similar in both seasons, while in the vineyards it varied, which can be explained by the cultivation practices that took place mainly in the spring, reducing the density of mites.
In the present study 59 species of Oribatida were found, and 57 of them were reported from the vineyards. Similar number of oribatid species was found in Italy (51 species; Nannelli and Simoni 2002) and Azerbaijan (61 species; Samedov et al. 1987), but was lower in Japan (24 species; Suzuki 1979) and India (16 species; Acharya and Basu 2014). Agricultural treatments of vineyards reduced the species richness of Oribatida, which is consistent with the conclusions of Meseguer Cervera (2014) and García-Parra (2015) who made similar observations on other arthropod groups in the same vineyards.
In the vineyards, Oribatula excavata dominated, followed by Minunthozetes quadriareatus and Passalozetes africanus. Oribatula excavata is a common species in Europe that is considered eurytopic, little sensitive to humidity conditions or organic matter content, and often found in cultivated fields (Pérez-Íñigo 1993) and in meadows (Weigmann 2006). Minunthozetes quadriareatus has a Mediterranean occidental distribution and is found in cultivated fields (Mingue et al. 1986). It is considered xerophilous, with preferences to cultivated soils (Pérez-Íñigo 1993), what explains its higher abundance in the vineyards than in the control. Similarly, Passalozetes africanus is a xerophilous species that is characteristic for very dry Mediterranean environment. It is tolerant of high temperatures and found in the areas without vegetation (Pérez-Íńigo 1993), which explains its highest dominance in the area between the vine rows, driven by a tractor, that was devoid of vegetation.
In the control, most abundant was Oppiella subpectinata, followed by Eremulus flagellifer. The former species is considered eurytopic, found in various habitats, but preferring higher content of organic matter (Weigmann 2006). This is why it was very abundant in the natural, undisturbed habitat, and in the vineyards was found exclusively in the areas between the vines, where the content of organic matter was higher than in other microhabitats. Eremulus flagellifer is a cosmopolitan species that requires a high humidity for development (Pérez-Íñigo 1997). This may explain why it was found only in the autumn, when precipitation was five-fold higher than in the spring.
In the vineyards in Italy (Nannelli and Simoni 2002), the most abundant was Tectocepheus velatus, which is eurytopic and easily colonizes new habitats. In the present study it also occurred in most plots, but in low numbers. In the vineyards in Japan, Scheloribates latipes was recorded in high number (Suzuki 1979), but in the present study it occurred in low numbers, while more common and more abundant was congeneric S. barbatulus. The presence of the species from genera Scheloribates and Protoribates in the vineyards is interesting, because they possibly play some role in control of pathogen fungi (Nakamura et al. 1991; Enami and Nakamura 1996).
A new species for Spain, Podoribates longipes, was found only in the control. It has a Holarctic distribution and was found in meadows (Migliorini et al. 2003), including salty ones (Weigmann 2006). Steganacarus boulfekhari was found only at the border of organic vineyard and has been known exclusively from Algeria, where it was recorded from many localities, mainly under different pine species (Pinus halepensis Mill., P. pinaster Aiton, P. canariensis C. Smith, P. radiata D. Don), oaks (Quercus faginea Lam.), eucalyptus (Eucalyptus L'Hér.), in evergreen bushes with Erica arborea L. and Pistacia L., and in orchards (Niedbała 2008).
In conclusion we can say that the oribatid mites do not benefit from organic cultivation of vineyards, probably because they are tolerant to the herbicides used in conventional vineyards, but are sensitive to the mechanical works on soil, which are more numerous in organic vineyards than in conventional ones. This study also supports the opinion (e.g. Duflot et al. 2015 and included references) that natural habitats in the agricultural landscape are important zones, increasing its total species diversity.
We are very grateful to Prof. Juan Carlos Iturrondobeitia (University of the Basque Country) for his invaluable comments and very helpful discussions on earlier versions of this paper.
Acharya S., Basu P. 2014. Diversity and richness of soil oribatid mites (Acarina: Arachnida: Arthropada) in grape orchads, Nashik, Maharashtra. Int. J. Res. Stud. Biosci., 2(7): 23-27.
Adán A., Vińuela E., Jacas J.-F. 1991. Effects of agricultural impact on soil inhabiting oribatid (Acari: Oribatida) communities. In: Dusbabek F., Bukva V. (Eds.). Modem Acarology. Academia, Prague and SPB Academic Publishing bv, The Hague, Vol. 1. p. 403-409.
Badejo M.-A., De Aquino A.-M., De-Polli H., M.-E. Fernandes Correia 2004. Response of soil mites to organic cultivation in an ultisol in southeast Brazil. Exp. Appl. Acarol. 34: 345-365. doi:10.1007/s10493-004-0201-y ![]()
Barbér Vallés A., Moity Martín N. 2009. Nueva población de Ononis rentonarensis (Fabaceae) en La Marina Alta (Alicante): implicaciones taxonómicas, fitogeográficas y geobotánicas. Flora Montiberica 44: 80-91.
Behan-Pelletier V.-M. 1999. Oribatid mite biodiversity in agroecosystems: role for bioindication. Agric. Ecosyst. Environ. 74: 411-423. doi:10.1016/S0167-8809(99)00046-8 ![]()
Bengtsson J., Ahnström J., Weibull A.-C. 2005. The effects of organic agriculture on biodiversity and density: a meta-analysis. J. Appl. Ecol. 42: 261-269. doi:10.1111/j.1365-2664.2005.01005.x ![]()
Berthet P., Gerard G. 1965. A statistical study of microdistribution of Oribatei (Acari). Part I. The distribution pattern. Oikos 16: 214-227.
Bettiol W., Ghini R., Haddad Galvăo J.-A., Vieira Ligo M.-A., de Carvalho MineiroJ.L. 2002. Soil organisms in organic and conventional cropping systems. Sci. Agr. 59(3): 565-572.
Doles J.-L., Zimmerman R.-J., John C. 2001. Soil microarthropod community structure and dynamics in organic and conventionally managed apple orchards in Western Colorado, USA. Appl. Soil Ecol. 18: 83-96. doi:10.1016/S0929-1393(01)00143-3 ![]()
Domingo J., Laborda R., Rodrigo E., Sánchez A., Bertomeu S., Xamani P. 2014. Biomoscatell de El Poble Nou de Benitatxell: conversión agroecológica y análisis de la emisión gases de efecto invernadero y consumos energéticos. III Remedia Workshop. 10-11 April, València.
Duflot R., Aviron S., Ernoult A., Fahrig L., Burel F. 2015. Reconsidering the role of 'semi-natural habitat' in agricultural landscape biodiversity: a case study. Ecol. Res. 30: 75-83. doi:10.1007/s11284-014-1211-9 ![]()
Enami Y., Nakamura Y. 1996. Influence of Scheloribates azumaensis (Acari: Oribatida) on Rhizoctonia solani, the cause of radish root rot. Pedobiologia 40: 251-254.
Fabeiro C., Gonzálvez V., Uranga J. 2007. Organic viticulture and wine making standards in Spain: state of the art. http://www.agroecologia.net/recursos/proyectos/informes/2004-07/organic%20viticulture%20%20wine%20in%20Spain%20englishcheck_16Jan07.pdf ![]() (accessed in June 2016)
(accessed in June 2016)
Fuangarworn M., Lekprayoon C., Pradatsundarasar A.-O. 2002. The short-term effects of atrazine herbicide on soil oribatid mites in a mango orchard.. The Natural History Journal of Chulalongkorn University 2(2): 1-5.
García-Parra I. 2015. Diversidad de ácaros edáficos (Acari: Mesostigmata) en cultivo de vińa en el Poble Nou de Benitatxell, Alicante. Unpublished MSc thesis, Universitat Politècnica de València, pp. 59.
Hill M.-O., Gauch H.-G. 1980. Detrended correspondence analysis, an improved ordination technique. Vegetatio 42: 47-58.
Johann L., Horn T.-B., Carvalho G.-S., Ferla N.-J. 2014. Diversity of mites (Acari) in vineyard agroecosystems (Vitis vinifera) in two viticultural regions of Rio Grande do Sul state, Brazil. Acarologia 54(2): 137-154. doi:10.1051/acarologia/20142122 ![]()
Jörger V. 1991. The influence of soil cultivation methods on the edaphic fauna, and especially the Gamasina (Mesostigmata), in two southern German vineyards with different cultural treatments. In: Schuster R. et al. (Eds.). The Acari. Springer Science+Business Media Dordrecht. p. 483-484. doi:10.1007/978-94-011-3102-5_51 ![]()
Lebrun Ph. 1977. Incidences écologiques des pesticides sur la faune du sol. Pédologie 27(1): 67-91.
Leps J., Smilauer P. 2003. Multivariate Analysis of Ecological Data Using CANOCO. Cambridge University Press, Cambridge. pp. 269.
Łomnicki A. 2010. Wprowadzenie do statystyki dla przyrodników. PWN, Warszawa. pp. 281.
Mäder P., Fliessbach A., Dubois D., Gunst L., Fried P., Niggli U. 2002. Soil fertility and biodiversity in organic farming. Science 296: 1694-1697.
MAGRAMA, Spanish Ministry of Agriculture, Food and the Environment, 2015; http://www.magrama.gob.es/es/ (accessed in June 2016).
Menhinick E.-F. 1962. Comparison of invertebrate populations of soil and litter of mowed grasslands in areas treated and untreated with pesticides. Ecology 43(3): 556-561.
Meseguer Cervera E. 2014. Comparació de l'abundància i diversitat d'artropodes entre parcelles de vinya ecologica i convencional i la influència del paisatge a El Poble Nou de Benitatxell. Unpublished Bachelor thesis, Universitat Politècnica de València, pp.75.
Migliorini M., Fanciulli P.-P., Bernini F. 2003. Comparative analysis of two edaphic zoocoenoses (Acari Oribatida; Hexapoda; Collembola) in the area of Orio al Serio Airport (Bergamo, northern Italy). Pedobiologia 47: 9-18. doi:10.1078/0031-4056-00164 ![]()
Mingue M.-E., Subías L.-S., Rui E. 1986. Dos nuevas especies de Oribátidos (Acari, Oribatida) de suelos cultivados de Espańa central. Boletín Asoc. esp. Entom. 10: 21-29.
Nakamura Y., Itakura J., Matzuzaki I. 1991. Mycophagous meso soil animals from cropfields in Fukushima Pref. Edaphologia 45: 49-54.
Nannelli R., Simoni S. 2002. Influence of different types of grass mulching on the communities of Oribatid mites in the vineyards. In: Bernini F., Nannelli R., Nuzzaci G., de Lillo E. (Eds.). Acarid Phylogeny and Evolution: Adaptation in Mites and Ticks, Kluwer Academic Publishers. p. 363-371. doi:10.1007/978-94-017-0611-7_37 ![]()
Niedbała W. 1980. Mechowce – roztocze ekosystemów lądowych. PWN Warszawa. pp. 255.
Niedbała W. 2008. Ptyctimous mites (Acari, Oribatida) of Poland. Fauna Poloniae, Natura optima dux Foundation 3, pp. 242.
Odum E.-P. 1982. Podstawy ekologii. PWRiL Warszawa pp. 661.
OIV 2016. http://www.oiv.int/public/medias/5029/world-vitiviniculture-situation-2016.pdf ![]() . (accessed in October 2017).
. (accessed in October 2017).
Pérez-Íñigo C. 1993. Acari, Oribatei, Poronota. In: Ramos M.-A. (Ed.). Fauna Ibérica, vol. 3. Museo Nacional de Ciencias Naturales CSIC, Madrid. pp. 320.
Pérez-Íñigo C. 1997. Acari. Oribatei. Gymnonota I.. In: Ramos M.-A. (Ed.), Fauna Ibérica, vol. 9. Museo Nacional de Ciencias Naturales CSIC, Madrid. pp. 373.
Piernik A. 2008. Metody numeryczne w ekologii na przykładzie zastosowań pakietu MVSP do analiz roślinności. Wyd. Nauk UMK, Toruń. pp. 103.
Samedov N.-G., Rasulova Z.-K., Kulagina I.-N., Loginova N.-G., Bababekova L.-A. 1987. The role of soil organisms in the increase of biological productivity in vineyards. In: Striganova B.-R. (Ed.), Soil fauna and soil fertility, Procceedings of the 9th International Colloquium on Soil Zoology, Moscow. p. 460-464.
Statistics National Institute: Agrarian Census 2009. http://ec.europa.eu/eurostat/statistics-explained/index.php/Agricultural_census_in_Spain ![]() . (accessed in June 2016).
. (accessed in June 2016).
Subías L.-S. 2004, 2015. Listado sistemático, sinonímico y biogeográfico de los Ácaros Oribátidos (Acariformes: Oribatida) del Mundo (1758−2002). Graellsia 60: 3-305; http://escalera.bio.ucm.es/usuarios/bba/cont/docs/RO_1.pdf ![]() (accessed in March 2016). doi:10.3989/graellsia.2004.v60.iExtra.218
(accessed in March 2016). doi:10.3989/graellsia.2004.v60.iExtra.218 ![]()
Subías L.-S., Shtanchaeva U. 2012. Oribátidos ibéricos (Acari: Oribatida): Listado sistemático, incluyendo nuevas citas de una familia, cuatro géneros y veinticinco especies. Revista Ibérica de Aracnología 20: 85-103.
Suzuki K. 1979. Comparison of oribatid fauna between a vineyard without insecticides and fertilizers and its adjacent secondary forest. Memoirs of the Education Institute for Private Schools in Japan, 64: 125-137.
Van Leeuwen J.-P., Lehtinen T., Lair G.-J., Bloem J., Hemerik L., Ragnarsdóttir K.-V., Gísladóttir G., Newton J.-S., de Ruiter P.-C. 2015. An ecosystem approach to assess soil quality in organically and conventionally managed farms in Iceland and Austria -Soil 1: 83-101.
Walker G.-E. 1992. Root rot of grapevine rootlings in South Australia caused by Rhizoctonia solani. Australas. Plant Path. 21(2): 58-60.
Walter D.-E., Proctor H. 1999. Mites. Ecology, evolution and behavior. University of New South Wales Press and CAB International, 1-322.
Weigmann G. 2006. Hornmilben (Oribatida). Die Tierwelt Deutschlands. Vol. 76, Goecke and Evers Keltern. Pp. 520.
Winter J.-P., Voroney R.-P., Ainsworth D.-A. 1990. Soil microarthropods in long-term no tillage and conventional tillage corn production. Can. J. Soil Sci. 70: 641-653. doi:10.4141/cjss90-067 ![]()



2016-10-16
Date accepted:
2018-05-24
Date published:
2018-09-28
Edited by:
Sidorchuk, Ekaterina

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Seniczak, Anna; Seniczak, Stanisław; García-Parra, Ivan; Ferragut, Francisco; Xamaní, Pilar; Graczyk, Radomir; Messeguer, Enric; Laborda, Rafael and Rodrigo, Eugenia
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)