




Use of predatory mites in commercial biocontrol: current status and future prospects
Knapp, Markus1 ; van Houten, Yvonne2 ; van Baal, Elmer3 and Groot, Thomas4
1✉ Koppert Biological Systems, P.O. Box 155, 2650 AD Berkel en Rodenrijs, the Netherlands.
2Koppert Biological Systems, P.O. Box 155, 2650 AD Berkel en Rodenrijs, the Netherlands.
3Koppert Biological Systems, P.O. Box 155, 2650 AD Berkel en Rodenrijs, the Netherlands.
4Koppert Biological Systems, P.O. Box 155, 2650 AD Berkel en Rodenrijs, the Netherlands.
2018 - Volume: 58 Issue: Suppl pages: 72-82
https://doi.org/10.24349/acarologia/20184275Keywords
Abstract
Biological control is a key ecosystem service and an underlying pillar of integrated pest management (IPM) (Naranjo et al. 2015). In many parts of the world, augmentative biological control of insect and mite pests is common practice as part of the IPM strategy in greenhouse cultivation of vegetables, fruits and ornamentals and 80% of the revenue created by commercial biological control is created in protected cultivation (Pilkington et al. 2010). Predatory mites, mainly from the family Phytoseiidae play the leading role among the biocontrol agents used. This paper will (1) give an overview on the importance of predatory mites in biocontrol; (2) show research needs to improve biological control, mainly with regard to soil-inhabiting predatory mites and present some preliminary experiments on this; (3) describe the potential of predatory mites in the control of poultry red mite, Dermanyssus gallinae (DeGeer).
The market for arthropod biological control agents has been growing with a rate of around 15% per year in the last decades (Ravensberg 2015). In 1997, the worldwide turnover of natural enemies was only about US 30 million (Bolckmans 1999), this increased to about US 50 million in 2000 (van Lenteren 2007) and to about US 400 million in 2010 (van Lenteren 2012); in 2014 it was about US 600 million.
The total number of arthropod species commercially available for augmentative biological control worldwide was about 125 in 2000 (van Lenteren 2000). In the next decade about 100 species were added. Of the 219 species listed by van Lenteren (2012) more than half were Hymenoptera (52.2%) followed by Acari (13.1%), Coleoptera (12.2%) and Heteroptera (8.3%). The 25 most frequently used species make up more than 90% of the turnover. The vast majority of them is used to control whiteflies, thrips, spider mites and aphids (van Lenteren 2012). This is also clearly to see in the ranking of the most important natural enemies. In the 1990s, 25% of the market was taken by the whitefly parasitoid Encarsia formosa Gahan, followed by the spider mite predator Phytoseiulus persimilis Athias-Henriot (12%) and the thrips predator Neoseiulus cucumeris (Oudemans) (12%) (Bolckmans 1999). This situation has changed in the last years. Currently more than 60% of the market is accounted for by phytoseiid predatory mites with Amblyseius swirskii (Athias-Henriot), P. persimilis, Neoseiulus californicus (McGregor) and N. cucumeris as the major species (Table 1). One of the major drivers of this change was the introduction of A. swirskii into the market in 2006. This species can control thrips and whiteflies simultaneously. It was the first predatory mite available that was effective against whiteflies (Nomikou et al. 2001), which until then were mainly controlled by parasitoids. The major advantages of A. swirskii compared to parasitoids are (1) it has a generalist feeding habit and can establish and multiply on pollen and other prey and therefore be introduced preventively, before the target pests are present in many crops; (2) it is easy to produce because it is small, multiplies quickly and can be reared on stored product mites, and (3) it can control several pests simultaneously. These characteristics made it possible that A. swirskii developed into the most widely used biocontrol agent in protected cultivation worldwide within only 10 years (Calvo et al. 2015).

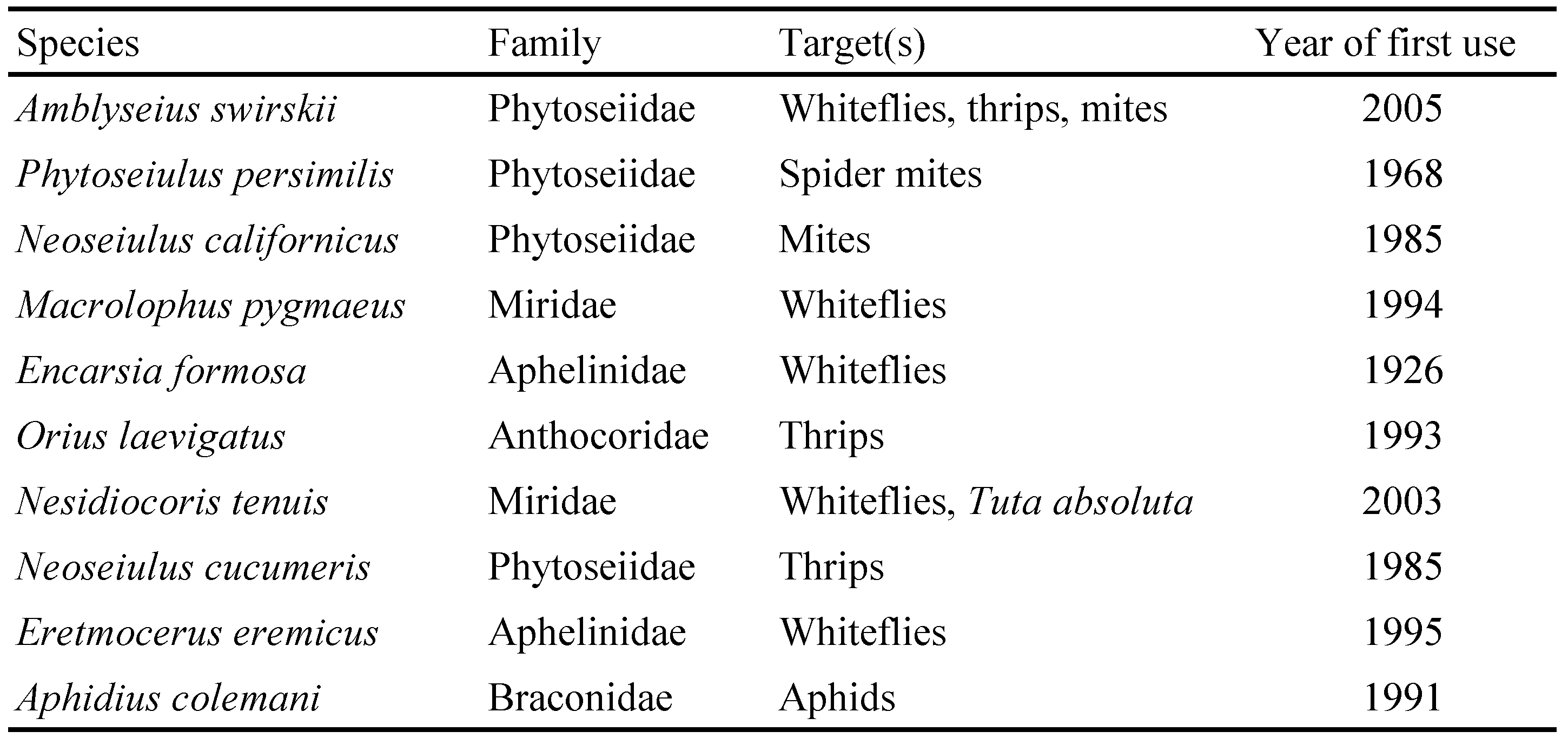
The majority of the predatory mites commercially available in Europe are belonging to the Phytoseiidae, followed by Laelapidae and one species each of Macrochelidae and Cheyletidae (Table 2). The rise of phytoseiids in importance for biological control in turn increased the interest in applied aspects of phytoseiid biology and a growing number of publications on, for instance influence of prey abundance and diversity on performance (Messelink et al. 2008), intra-guild interaction (Abad-Moyano et al. 2010; Buitenhuis et al. 2010), influence of host plants (Beard and Walter 2001; van Houten et al. 2013; Buitenhuis et al. 2014) or on the evaluation of biocontrol using phytoseiids in commercial crops (Gacheri et al. 2015; Sampson and Kirk 2016) help biocontrol agent producers and biocontrol practitioners to understand the behaviour of phytoseiids and to further improve biological control.

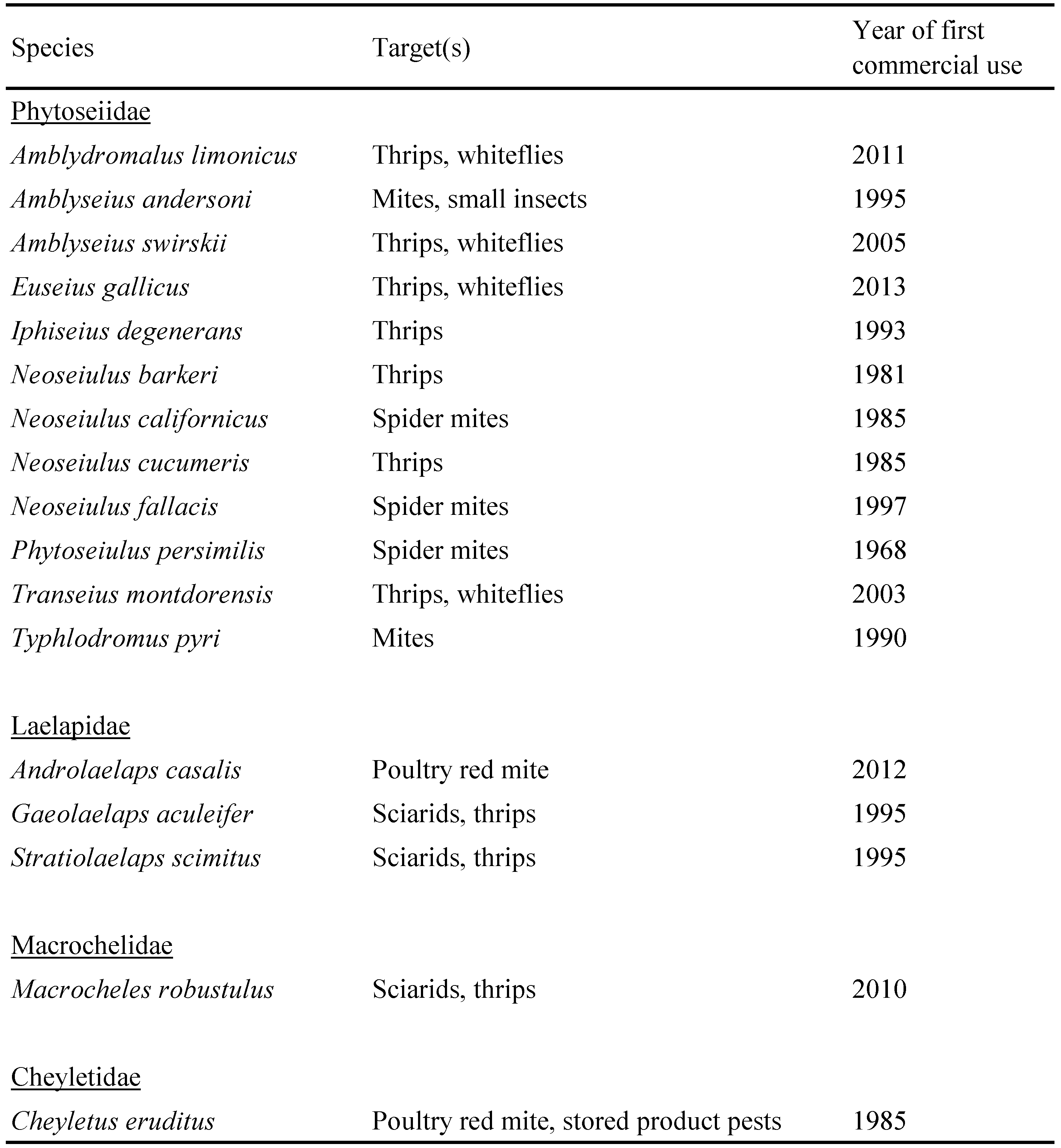
Compared to the phytoseiids, soil-dwelling predatory mites so far play a minor role in biological control. The four species currently on the market, Gaeolaelaps aculeifer (Canestrini), Gaeolaelaps gillespiei Beaulieu, Stratiolaelaps scimitus (Womersley) (Laelapidae) and Macrocheles robustulus (Berlese) (Macrochelidae) probably account for less than 2% of the turnover with natural enemies. This group is also much less researched than the Phytoseiidae; the number of records in the Web of Science core collection is 2109 for Phytoseiidae, 229 for Laelapidae and 109 for Macrochelidae (check date 16 June 2016). The biology and use in biological control of these soil-dwelling species was recently reviewed (Moreira and Moraes 2015; De Azevedo et al. 2015). All of them can feed on a wide range of prey, however few studies have been conducted to validate their effectivity in field experiments and results are not consistent (see e.g. Manners et al. 2013; Pozzebon et al. 2015). Despite this, a significant number of greenhouse growers regularly apply soil-dwelling predatory mites to control sciarid larvae and thrips. One of the challenges in semi-field and field studies is the quantitative assessment of the density of the soil-dwelling predatory mites released. Usually Berlese-Tullgren funnels are used for this purpose (Freire et al. 2007; Grosman et al. 2011), however the size and design of these funnels can vary widely and data on their extraction efficiency are lacking. Therefore, we have conducted some preliminary experiments on the extraction efficiency of a commercially available Berlese-Tullgren funnel (see below).
Berlese-Tullgren funnels are widely used to extract arthropods from soil or vegetation samples (Edwards 1991, André et al. 2002). However, they have mostly been used for ecological or taxonomic studies and their extraction efficiency for specific soil arthropods is very variable. There are very few studies investigating extraction efficiency by recapture of known numbers of specific arthropods added to defaunated soil or other medium (Brady 1969; André et al. 2002).
For our experiments we used a commercially available Berlese-Tullgren apparatus consisting of 12 funnels (Burkard Scientific, Uxbridge, UK). This apparatus is supplied with 25 W light bulbs by the manufacturer. As the extractions took very long in preliminary trials, we also tested 40 W bulbs in some experiments. The removable upper part of the funnels is equipped with a 2 mm mesh at the bottom. To reduce the amount of debris falling into the sampling vials we added a finer mesh (800 µm) and put 10 g of buckwheat husks on top of this mesh before adding potting soil. The collection vials fixed at the lower end of the funnels were filled with 70% alcohol. We used standard potting soil (50% coco peat, 15% white peat, 35% frozen black peat: pH 5.5, e.c. 1.0), which was sterilized to kill any potential pre-existing fauna and then re-moistened to a water content of approximately 70% (w/w). Experiments were conducted with the soil-dwelling predatory mite species M. robustulus and G. aculeifer. Both species were reared in the laboratory at 25 °C; M. robustulus was reared on Carpoglyphus lactis (Linnaeus) and G. aculeifer on Acarus siro (Linnaeus).
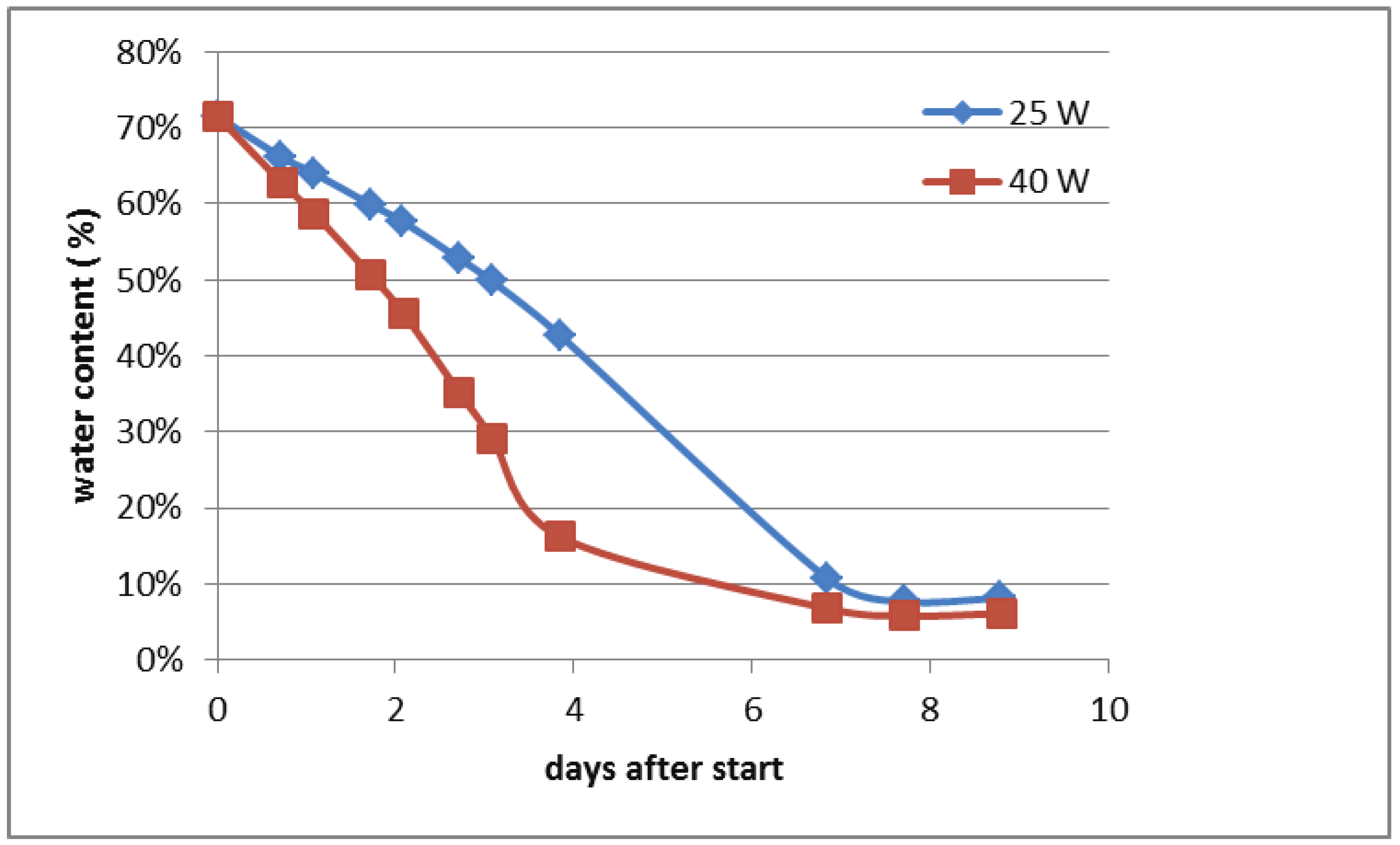
To determine the reduction in moisture content of the samples in the funnels with 25 W and 40 W bulbs, 500 mL of the potting soil (without mites) was filled in the removable upper part of funnels. This resulted in a layer of approximately 7.5 cm height. Thereafter the removable containers were placed into the Berlese apparatus equipped with 25 W and 40 W light bulbs, respectively. The moisture content was determined with a moisture analyser (Sartorius MA 160, Sartorius Lab Instruments, Göttingen, Germany) immediately before the start of the experiment and thereafter by weighing the samples regularly for 9 days. At the end of the trial period the moisture content was determined again with the moisture analyser.
The water content was 71.5% when the experiment started and declined faster under the 40 W bulb than under the 25 W bulb. In both treatments the moisture content did not decline further after day 7 (Figure 1). The moisture content at the end of the trial was 8.1% and 9.2% with 40 W and 25 W, respectively.

Thirty adult predatory mites (80% females) were added to 500 mL moist potting soil and carefully mixed in a plastic bowl. In the treatments with C. lactis several thousand C. lactis were added as prey for the M. robustulus. This was done because samples from the field usually also contain prey for the predatory mites. Thereafter the samples were put on top of the buckwheat in the removable containers of the Berlese-Tullgren apparatus equipped with 40 W lights. The containers were placed into the apparatus and the lights were switched on after 10 minutes. Three containers (replications) were prepared per treatment.
Without food 63% of the mites were extracted after 7 days. No more mites came out of the sample during the next two days. When C. lactis was provided as food, more mites came out of the samples than initially were put in. On day 5, an average of 52.7 mites had been collected in the collection vials; 50% of them were immatures (Figure 2).


The setup was similar to the experiment with M. robustulus with the following exceptions. The Berlese-Tullgren funnel was equipped with 25 W lamps; A. siro was used as prey mite instead of C. lactis and 20 adult G. aculeifer (80% females) were used per funnel in the trial without prey mites, whereas 30 mites were used in the experiment with prey mites.
Without prey mites, 85% of the mites were recaptured, when A. siro was added G. aculeifer multiplied similarly to M. robustulus. More mites were extracted than introduced and the proportion of immatures was 49%. In contrast to the trial with M. robustulus also larvae were found in the extraction vials (Figure 3).


The most striking result of these experiments was that the predatory mites can multiply in the Berlese-Tullgren funnel if food is available. Moisture and temperature in certain areas of the sample seem to be conducive to the development of both predatory mite species. Egg to adult development of M. robustulus can be extremely fast; it takes about 2 days at 25-29 °C (Filipponi and Mosna 1968). Immature development of G. aculeifer is slower, however this species can develop from egg to adult in 8 to 10 days between 27 and 30 °C; the eggs hatch after around 3 days under these conditions (Amin et al. 2014).
Furthermore there are indications that the extraction of G. aculeifer is more efficient than the extraction of M. robustulus if no food is available.Some M. robustulus adults also left the soil sample by walking out of the top of the funnels. This was verified by putting double sided tape on the outer rim of the funnels, after we observed that prey mites were walking on the rim of the funnels. A few M. robustulus adults were found on this tape. As M. robustulus is phoretic on beetles (Glida et al., 2003; Philips, 2009), these mites might have walked to the upper surface in search a phoriont.
To our knowledge there is no other study investigating the re-capture of known numbers of released soil-dwelling predatory mites with Berlese-Tullgren funnels. Brady (1969) investigated the extraction efficiency of Caloglyphus berlesei (Michael), A. siro and Cheyletus trouessarti (Oudemans) from defaunated poultry litter; he recorded recovery rates of 86-97% for adults. Until a reliable extraction method has been developed, counts of soil-dwelling predatory mites extracted with Berlese-Tullgren funnels from comparative experiments with different species have to be interpreted with great care (see e.g. Messelink and van Holstein-Saj 2008; Grosman et al. 2011; Pijnakker et al. 2011).
The poultry red mite D. gallinae poses a serious threat to egg laying hens worldwide. Economic costs associated with control and production losses have been estimated at € 130 million per year for the European Union egg production industry. Dermanyssus gallinae has typically been controlled by using synthetic acaricides but resistance of the mites to acaricides and legislative restrictions on the use of acaricides make this approach more and more difficult (Sparango et al. 2014). The behaviour of the mites poses an additional challenge to their control. The majority of the life cycle is spent off the host where the mites hide in large numbers in secluded areas like cracks formed by timber joints or in metal profiles supporting the perches. Due to the difficulty to conduct experiments on life chicken few data are available on the life cycle of D. gallinae. Maurer and Baumgärtner (1992) found that immature development takes 7 days at 25 °C and calculated an intrinsic rate of increase (rm) of 0.120 at this temperature. Dermanyssus gallinae can develop between 5 and 40 °C but conditions for juvenile development are most favourable between 30 and 37 °C (Maurer and Baumgärtner 1992; Maurer and Hertzberg 2001).
Research on the control of D. gallinae with predatory mites has started later than research on using predatory mites to control plant pests. Maurer (1993) discovered that Cheyletus eruditus (Schrank) is associated with D. gallinae in Swiss poultry houses and can feed on its larvae. However, initial trials with this predatory mite species in poultry houses yielded disappointing results (Maurer and Hertzberg, 2001).
In a new effort to find biocontrol agents for poultry red mite Lesna et al. (2009) surveyed the mite fauna in starling nests and chicken houses in the Netherlands. The predators C. eruditus and A. casalis were frequently found both in starling nests and in chicken houses. Blattisocius keegani Fox was also common in chicken houses but not in starling nests. Small experiments in the laboratory confirmed that A. casalis and G. aculeifer predated on D. gallinae. The latter predator was rare in starling nests and absent in chicken houses, but is available commercially as a biocontrol agent. Later Ali et al. (2014) demonstrated that S. scimitus also predates the poultry red mite in laboratory trials.
In small laying hen cages A. casalis and S. scimitus reduced the D. gallinae population compared to untreated cages but A. casalis was more efficient than S. scimitus. However, both species were not able to completely eradicate the poultry red mites from the cages (Lesna et al. 2012).
The hunting behaviour of C. eruditus and A. casalis is different. Cheyletus eruditus is a “sit and wait”/ ambush predator. It rarely actively stalks its prey (Thind and Ford 2006). Androlaelaps casalis is hunting for prey very actively (Barker 1968). At 25.6 °C, 75-100% relative humidity and with Glycophagus domesticus (De Geer) as prey, development time of A. casalis from egg to adult is about 8 days (Barker 1968).
Although C. eruditus has been known to occur in poultry houses for long (Brady 1970) it has mainly been investigated as biological control agent of stored product mites (Zdarkova 1998). Maurer (1993) calculated an intrinsic rate of increase (rm) of 0.144, which is higher than the rm of D. gallinae (0.12, Maurer and Baumgärtner 1992) and a doubling time of 4.8 days for C. eruditus on D. gallinae at 25 °C. With Lepidoglyphus destructor (Schrank) as prey, egg to adult development of C. eruditus took 15.4 days, females laid 133 eggs (5.4 per day) and the rm was 0.178 (Barker 1991).
Releases of mass-reared C. eruditus as well as A. casalis in chicken houses have not led to a reliable control of D. gallinae (Maurer and Hertzberg 2001; Koppert unpubl. data). Therefore, a strategy was developed based on the combined use of these two predatory mites. In aviary systems, C. eruditus is released after cleaning the laying hen houses between two production cycles, one week before the new hens are introduced, by spreading the mite together with a carrier in the chicken houses. This strategy has been used in grain stores in the Czech Republic in the past (Zdarkova and Horak 1990) and gives the mites time to find a hiding place and to feed on any remaining poultry red mites. As the carrier is very attractive to the chicken and C. eruditus tends to remain in the carrier for some time most predatory mites are eaten by the hens quickly after spreading if the release is carried out when chicken are already present. The release of A. casalis starts two weeks after the laying hens have arrived and some dung and dirt has accumulated. Androlaelaps casalis is released by installing bottles containing predatory mites, prey mites and a carrier, close to typical hiding places of D. gallinae in the structure of the chicken house, or more humid places (dung accumulations). The predatory mites disperse through the perforated lid of the bottles. At the same time D. gallinae are also entering the bottles in search for hiding places and are subsequently fed on by the population of predatory mites inside. These bottles can sustain a large population of predatory mites for more than 8 weeks; thereafter they are replaced. This system takes into account observations of Lesna et al. (2012) who recorded that most poultry red mites were found higher up in the cage structure whereas the predatory mites A. casalis and S. scimitus were mainly found on the cage floor, and recommended to create artificial environments in which predatory mites can thrive and which can be positioned higher up in the structural elements of cage and aviary systems.
Biological control using leaf-inhabiting phytoseiid predatory mites has been booming during the last decades. This was made possible through close collaboration between researchers and biocontrol companies. Nevertheless, there are many challenges remaining, for instance developing better systems for ornamentals where tolerance levels for pests are very low, possibly by using providing alternative food sources for the predators (see e.g. Buitenhuis et al. 2015; Delisle et al. 2015; Janssen and Sabelis 2015) or finding generalist predatory mites that can successfully control pests on tomatoes (see e.g. van Houten et al. 2013).
The potential for the use of soil-dwelling predatory mites could be as high as for leaf-inhabiting predatory mites. Many species of soil-dwelling predatory mites are known to feed on important plant pests (Carrillo et al. 2015). However, very little is known about them besides that they prey on a certain pest species and to realize their potential we need collaborative efforts to develop further understanding of the biology and ecology of these mites and their interaction with other soil organisms. We have shown that currently even reliable methods to quantify the density of soil-dwelling predatory mites are lacking. Development of such a method would be an important first step.
Control of poultry red mite has received more and more attention during the last few years. Initial results with A. casalis and C. eruditus are promising in aviary systems but satisfactory control will only be possible in combination with other measures.
Generally, mites have a huge potential in biological control. There small size and fast population development makes them relatively easy to rear and to transport to the end users. Further improvements of mass-rearing and introduction techniques will make biological control using predatory mites even more robust and reliable as well as more affordable for growers.
We thank Alma van der Heiden for her assistance with the soil mite extraction experiments.
Abad-Moyano R., Urbaneja A., Schausberger P. 2010. Intraguild interactions between Euseius stipulatus and the candidate biocontrol agents of Tetranychus urticae in Spanish clementine orchards: Phytoseiulus persimilis and Neoseiulus californicus. Exp. Appl. Acarol., 50: 23-34. doi:10.1007/s10493-009-9278-7 ![]()
Ali W., George D.R., Shiel R.S., Sparagano O.A., Guy J.H. 2012. Laboratory screening of potential predators of the poultry red mite (Dermanyssus gallinae) and assessment of Hypoaspis miles performance under varying biotic and abiotic conditions. Vet. Parasitol., 187: 341-344. doi:10.1016/j.vetpar.2012.01.014 ![]()
Amin M.R., Khanjani M., Zahiri B. 2014. Estimating development of preimaginals and fecundity of Gaeolaelaps aculeifer (Acari: Laelapidae) feeding on Rhizoglyphus echinopus (Acari: Acaridae) at constant temperatures. J. Crop Prot., 3: 581-587.
André H.M., Ducarme X., Lebrun P. 2002. Soil biodiversity: myth, reality or conning?. Oikos, 96: 3-24. doi:10.1034/j.1600-0706.2002.11216.x ![]()
Barker P.S. 1968. Bionomics of Androlaelaps casalis (Berlese) (Acarina: Laelapidae) a predator of mite pests of stored cereals. Can. J. Zool., 46: 1099-1102. doi:10.1139/z68-157 ![]()
Barker P.S. 1991. Bionomics of Cheyletus eruditus (Schrank) (Acarina: Cheyletidae), a predator of Lepidoglyphus destructor (Schrank) Acarina: Glycyphagidae), at three constant temperatures. Can. J. Zool., 69: 2321-2325. doi:10.1139/z91-326 ![]()
Beard J.J., Walter G.H. 2001. Host plant specificity in several species of generalist mite predators. Ecol. Entomol., 26: 562-570. doi:10.1046/j.1365-2311.2001.00367.x ![]()
Bolckmans K.J.F, 1999. Commercial aspects of biological pest control. In: Albajes R., Gullino M.L., van Lenteren J.C., Elad Y. (Eds.). Integrated Pest and Disease Management in Greenhouse Crops; Dordrecht: Kluwer. p. 310-318. doi:10.1007/0-306-47585-5_22 ![]()
Brady J. 1969. Some physical gradients set up in Tullgren funnels during the extraction of mites from poultry litter. J. Appl. Ecol., 6: 391-402. doi:10.2307/2401506 ![]()
Brady, J. 1970. The mites of poultry litter: Observations on the bionomics of common species, with a species list for England and Wales. J. Appl. Ecol., 7: 331-348. doi:10.2307/2401384 ![]()
Buitenhuis R., Shipp L., Scott-Dupree C. 2010. Intra-guild vs extra-guild prey: effect on predator fitness and preference of Amblyseius swirskii (Athias- Henriot) and Neoseiulus cucumeris (Oudemans) (Acari: Phytoseiidae). Bull. Entomol. Res., 100: 167-173. doi:10.1017/S0007485309006944 ![]()
Buitenhuis R., Shipp L., Scott-Dupree C., Brommit A., Lee W. 2014. Host plant effects on the behaviour and performance of Amblyseius swirskii (Acari: Phytoseiidae). Exp. Appl. Acarol., 62: 171-180. doi:10.1007/s10493-013-9735-1 ![]()
Buitenhuis R., Murphy G., Shipp L., Scott-Dupree C. 2015. Amblyseius swirskii in greenhouse production systems: a floricultural perspective. Exp. Appl. Acarol. 65: 451-464. doi:10.1007/s10493-014-9869-9 ![]()
Calvo F.J., Knapp M., van Houten Y.M., Hoogerbrugge H., Belda J.E. 2015. Amblyseius swirskii: What made this predatory mite such a successful biocontrol agent?. Exp. Appl. Acarol., 65: 419-433. doi:10.1007/s10493-014-9873-0 ![]()
Carrillo D., de Moraes G.J., Pena J.E. (Eds.) 2015. Prospects for biological control of plant feeding mites and other harmful organisms. Progress in Biological Control 19; Cham: Springer. xiv + 328pp.
De Azevedo L.H., Emberson R.M., Esteca F. de C.N., de Moraes G.J. 2015. Macrochelid mites (Mesostigmata: Macrochelidae) as biological control agents. In: Carrillo D., de Moraes G.J., Pena J.E. (Eds.). Prospects for biological control of plant feeding mites and other harmful organisms. Progress in Biological Control 19; Cham: Springer. p. 103-132.
Delisle J.F., Shipp L., Brodeur J. 2015. Apple pollen as a supplemental food source for the control of western flower thrips by two predatory mites, Amblyseius swirskii and Neoseiulus cucumeris (Acari: Phytoseiidae), on potted chrysanthemum. Exp. Appl. Acarol., 65: 495-509. doi:10.1007/s10493-014-9863-2 ![]()
Edwards C.A. 1991. Methods for assessing populations of soil-inhabiting invertebrates: The assessment of populations of soil-inhabiting invertebrates. Agric. Ecosys. Environ., 34: 145-176. doi:10.1016/0167-8809(91)90102-4 ![]()
Filipponi A., Mosna B., 1968. Influenza di fattori ecologici e genetici sulla natalita e mortalita di Macrocheles robustulus (Berlese, 1904) Ann. Ist. Super. Sanita, 4: 551-571.
Freire R.A.P., Moraes G.J., Silva E.S., Vaz A.C., Campos Castilho R. 2007. Biological control of Bradysia matogrossensis (Diptera: Sciaridae) in mushroom cultivation with predatory mites. Exp. Appl. Acarol., 42: 87-93. doi:10.1007/s10493-007-9075-0 ![]()
Gacheri C., Kigen T., Sigsgaard L. 2015. Hot-spot application of biocontrol agents to replace pesticides in large scale commercial rose farms in Kenya. BioControl, 60: 795-803.
Glida H., Bertrand M., Peyrusse V. 2003. A limiting factor in the abundance of predatory phoretic mites (Acari: Macrochelidae): the seasonal abundance of their phorionts (dung beetles) in southern France. Can. J. Zool., 81: 2066-2072. doi:10.1139/z03-199 ![]()
Grosman A., Messelink G.J., de Groot E. 2011. Combined use of a mulch layer and the soil-dwelling predatory mite Macrocheles robustulus (Berlese) enhance the biological control of sciarids in potted plants. IOBC/wprs Bull., 68: 51-54.
Janssen A., Sabelis M.W. 2015. Alternative food and biological control by generalist predatory mites: the case of Amblyseius swirskii. Exp. Appl. Acarol., 65: 413-418. doi:10.1007/s10493-015-9901-8 ![]()
Lesna I., Wolfs P., Faraji F., Roy L., Komdeur J., Sabelis M.W. 2009. Candidate predators for biological control of the poultry red mite Dermanyssus gallinae. Exp. Appl. Acarol., 48: 63-80. doi:10.1007/s10493-009-9239-1 ![]()
Lesna I., Sabelis M.W., van Niekerk T.G., Komdeur J. 2012. Laboratory tests for controlling poultry red mites (Dermanyssus gallinae) with predatory mites in small 'laying hen' cages. Exp. Appl. Acarol., 58: 371-383. doi:10.1007/s10493-012-9596-z ![]()
Manners A.G., Dembowski B.R., Healey M.A. 2013. Biological control of western flower thrips, Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae), in gerberas, chrysanthemums and roses. Austral. J. Entomol., 52: 246-258. doi:10.1111/aen.12020 ![]()
Maurer V. 1993. The dynamics of Dermanyssus gallinae (Acari: Dermanyssidae) populations interacting with laying hens and the predatory mite Cheyletus eruditus (Acari: Cheyletidae) [PhD Thesis]. Zürich: Swiss Federal Institute of Technology.
Maurer V., Baumgärtner J. 1992. Temperature influence on the life table statistics of the chicken mite Dermanyssus gallinae (Acari: Dermanyssidae). Exp. Appl. Acarol., 15: 27-40. doi:10.1007/BF01193965 ![]()
Maurer V., Hertzberg H. 2001. Ökologische Legehennenhaltung: Was tun gegen die kleinen Vampire?. DGS Magazin Woche 40/2001: 49-52.
Messelink G.J., van Holstein-Saj R. 2008. Improving thrips control by the soil-dwelling predatory mite Macrocheles robustulus (Berlese). IOBC/wprs Bull., 32: 135-138.
Messelink G.J., van Maanen R., van Steenpaal S.E.F., Janssen, A. 2008. Biological control of thrips and whiteflies by a shared predator. Two pests are better than one. Biol. Contr., 44: 372-379.
Moreira G.F., de Moraes G.J. 2015. The potential of free-living laelapid mites (Mesostigmata: Laelapidae) as biological control agents. In: Carrillo D., de Moraes G.J., Pena J.E. (Eds.). Prospects for biological control of plant feeding mites and other harmful organisms. Progress in Biological Control 19; Cham: Springer. p. 77-102. doi:10.1007/978-3-319-15042-0_3 ![]()
Naranjo S.E., Ellsworth P.C., Frisvold G.B. 2015. Economic value of biological control in integrated pest management of managed plant systems. Annu. Rev. Entomol. 60: 621-45. doi:10.1146/annurev-ento-010814-021005 ![]()
Nomikou M., Janssen A., Schraag R., Sabelis M.W. 2001. Phytoseiid predators as potential biological control agents for Bemisia tabaci. Exp. Appl. Acarol., 25: 271-291. doi:10.1023/A:1017976725685 ![]()
Philips J.R. 2009. The mite (Acarina) fauna of trogid beetles (Coleoptera: Trogidae). Int. J. Acarol. 35: 1-17. doi:10.1080/01647950802709843 ![]()
Pijnakker J., Leman A., Messelink G., Grosman A., van Holstein R. 2011. Bestrijding van rouwmuggen en oevervliegen. Rapport GTB-1090. Wageningen UR Glastuinbouw. Available from http://edepot.wur.nl/174724 ![]()
Pilkington L.J., Messelink G., van Lenteren J.C., Le Mottee K. 2010. "Protected Biological Control" – Biological pest management in the greenhouse industry. Biol. Contr., 52: 216-220.
Pozzebon A., Boaria A., Duso C. 2014. Single and combined releases of biological control agents against canopy-and soil-dwelling stages of Frankliniella occidentalis in cyclamen. BioControl, 60: 341-350.
Ravensberg W. 2015. Biocontrol as a business. 4th Symposium: Feeding Europe by reducing pesticide dependency, Brussels: European Parliament 19-11-2015. Available from http://www.ibma-global.org/upload/documents/4wravensbergproducerofbiologicalcontrolproductsepsymposium.pdf ![]()
Sampson C., Kirk W.D.J. 2016. Predatory mites double the economic injury level of Frankliniella occidentalis in strawberry. BioControl, 61: 661–669. doi:10.1007/s10526-016-9747-y ![]()
Sparagano O.A.E., George D.R., Harrington D.W.J. Giangaspero A. 2014. Significance and control of the poultry red mite, Dermanyssus gallinae. Annu. Rev. Entomol., 59: 447-66. doi:10.1146/annurev-ento-011613-162101 ![]()
Thind B.B., Ford H.L. 2006. Laboratory studies on the use of two new arenas to evaluate the impact of the predatory mites Blattisocius tarsalis and Cheyletus eruditus on residual populations of the stored product mite Acarus siro. Exp. Appl. Acarol., 38: 167-180. doi:10.1007/s10493-005-5829-8 ![]()
Van Houten Y.M., Glas J.J., Hoogerbrugge H., Rothe J., Bolckmans K.J.F., Simoni S., van Arkel J., Alba J.M., Kant M.R., Sabelis M.W. 2013. Herbivory-associated degradation of tomato trichomes and its impact on biological control of Aculops lycopersici. Exp. Appl. Acarol., 60: 127-138. doi:10.1007/s10493-012-9638-6 ![]()
Van Lenteren J.C. 2000. Success in biological control of arthropods by augmentation of natural enemies. In: Gurr G., Wratten S. (Eds.). Biological control: matters of success. Dordrecht: Kluwer. p. 77-103. doi:10.1007/978-94-011-4014-0_3 ![]()
Van Lenteren J.C. (ed.), 2007. Internet Book of Biological Control. 4th Edition, Wageningen: www.IOBC-Global.org. Available from http://www.iobc-global.org/download/IOBC_InternetBookBiCoVersion6Spring2012.pdf ![]()
Van Lenteren J.C. 2012. The state of commercial augmentative biological control: plenty of natural enemies, but a frustrating lack of uptake. BioControl, 57: 1-20.
Zdarkova, E. 1998. Biological control of storage mites by Cheyletus eruditus. Integr. Pest Manag. Rev. 3: 111-116 (1998) doi:10.1023/A:1009647814477 ![]()
Zdarkova E, Horak E. 1990. Preventive biological control of stored food mites in empty stores using Cheyletus eruditus (Schrank). Crop Prot., 9: 378-382. doi:10.1016/0261-2194(90)90011-U ![]()



2016-11-17
Date accepted:
2018-03-29
Date published:
2018-09-28
Edited by:
Kreiter, Serge

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Knapp, Markus; van Houten, Yvonne; van Baal, Elmer and Groot, Thomas
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)