




External morphology of postembrionic stages of Lutrilichus javanicus (Acariformes: Chirodiscidae) from Melogale moschata (Carnivora: Mustelidae) from Vietnam
Bochkov, Andre V.1
1✉ Zoological Institute, Russian Academy of Sciences, Universitetskaya emb. 1, 199034 St. Petersburg, Russia. and Museum of Zoology, University of Michigan, 1109 Geddes Ave., Ann Arbor, Michigan 8109, USA. and Tyumen State University, 10 Semakova Str., 625003, Tyumen, Russia.
2018 - Volume: 58 Issue: 4 pages: 763-772
https://doi.org/10.24349/acarologia/20184271ZooBank LSID: 9BA16FB7-71A7-4A07-8BBF-7083E5021B41
Keywords
Abstract
The family Chirodiscidae (Acariformes: Sarcoptoidea) includes approximately 230 species in 26 genera and four subfamilies (Bochkov 2010). These mites are permanent mono- or stenoxenous symbionts of mammals that live in the fur of their hosts. The tribe Schizocarpini (Labidocarpinae) includes three genera, Schizocarpus Trouessart from beavers (Rodentia: Castoridae) (60 species), Soricilichus Fain (3 species) from African shrews of the subfamily Crocidurinae (Soricomorpha: Soricidae), and Lutrilichus Fain (5 species) from small carnivores of the families Mustelidae and Viverridae (Carnivora) (Fain 1970, 1971, 1981; Fain et al. 1974; Bochkov et al. 2016). In mites of this tribe, the external morphology of immature instars strongly differs in male and female lines of development. Larvae are hexapode with well-developed legs III having a full set of setae typical for this stage in chirodiscids. Male protonymphs and tritonymphs also bear the full set of setae and have well-developed legs III and IV. All these preimaginal stages of the male line live independently. At the same time, female proto- and tritonymphs are sacciform, the number of their idiosomal setae is strongly reduced compared to those of the male line, and their legs III and IV are primordial or absent. They are apparently not able to live independently and are always found attached to males with the posterior end of the opisthosoma (Fain 1971).
The external morphology of the immature stages in chirodiscid genera Schizocarpus and Soricilichus was studied in detail by Dubinina (1964) and Bochkov et al. (2016), respectively. At the same time, the immature stages of Lutrilichus have never been studied specifically.
In this paper, the external morphology of all postembryonic stages of Lutrilichus javanicus Fain, 1970 collected from fur of Melogale moschata (Gray) (Carnivora: Mustelidae) in Vietnam is described . This mite species was previously known from a single female specimen (holotype, housed in the Natural History Museum, London, UK) from Melogale orientalis Blanford, from Java (Fain 1970; 1981).
The host of mite specimens used in the present study, the Chinese ferret badger, Melogale moschata, was collected by my colleague, A.V. Abramov (Zoological Institute of the Russian Academy of Sciences, Saint-Petersburg, Russia), in the zoological survey in northern Vietnam carried out by the Vietnam-Russia Tropical Center (Ho Chi Minh City — Moscow) in 2013. Mites were gathered by AB from the ethanol-preserved host with fine forceps under dissection microscope, placed in 96% ethanol, and then mounted in Hoyer’s medium according to standard methods (Evans 1992). Drawings were made with a Leica microscope equipped with differential interference contrast optics and a camera lucida.
In the description below, the idiosomal setation follows Griffiths et al. (1990) with modifications for coxal setae by Norton (1998), and leg setation follows Grandjean (1939). All measurements are in micrometers (μm) and were taken as follows: body length = length from the palpal apices line to the posterior margin of the body; idiosomal width = lateral width at the level of setae cp; length of dorsal shields = maximum length, measured along the median line of the shields; length of the posterior legs = length from the most proximal point of the trochanter to the apex of the tarsus, excluding the pretarsus.
Lutrilichus javanicus Fain, 1970 (Figures 1-7)
Lutrilichus javanicus Fain, 1970: 300, 1981: 43, figs. 3, 4; Fain et al. 1974: 944.
Material examined — 10 males, 10 females, 10 male larvae, 10 female larvae, 5 male protonymphs, 10 female protonymphs, 10 male tritonymphs, 10 female tritonymphs, and numerous specimens retained in ethanol (ZISP, AVB 17-1103-001) from the Chinese ferret badger Melogale moschata (Gray) (Carnivora: Mustelidae) (ZISP, AVA 13-169), VIETNAM: Son La Province, Phu Yen District, Suoi To Commune, Suoi Khang Village, ca. 10 km NW of Phu Yen, 21°20′13.2′′N, 104°36′29.7′′E, alt 1100 m, 31 May 2013, coll. A.V. Abramov. Mites are deposited in the Museum of Zoology, the University of Michigan, Ann Arbor, USA (UMMZ), and Zoological Institute of the Russian Academy of Sciences, Saint-Petersburg, Russia (ZISP).
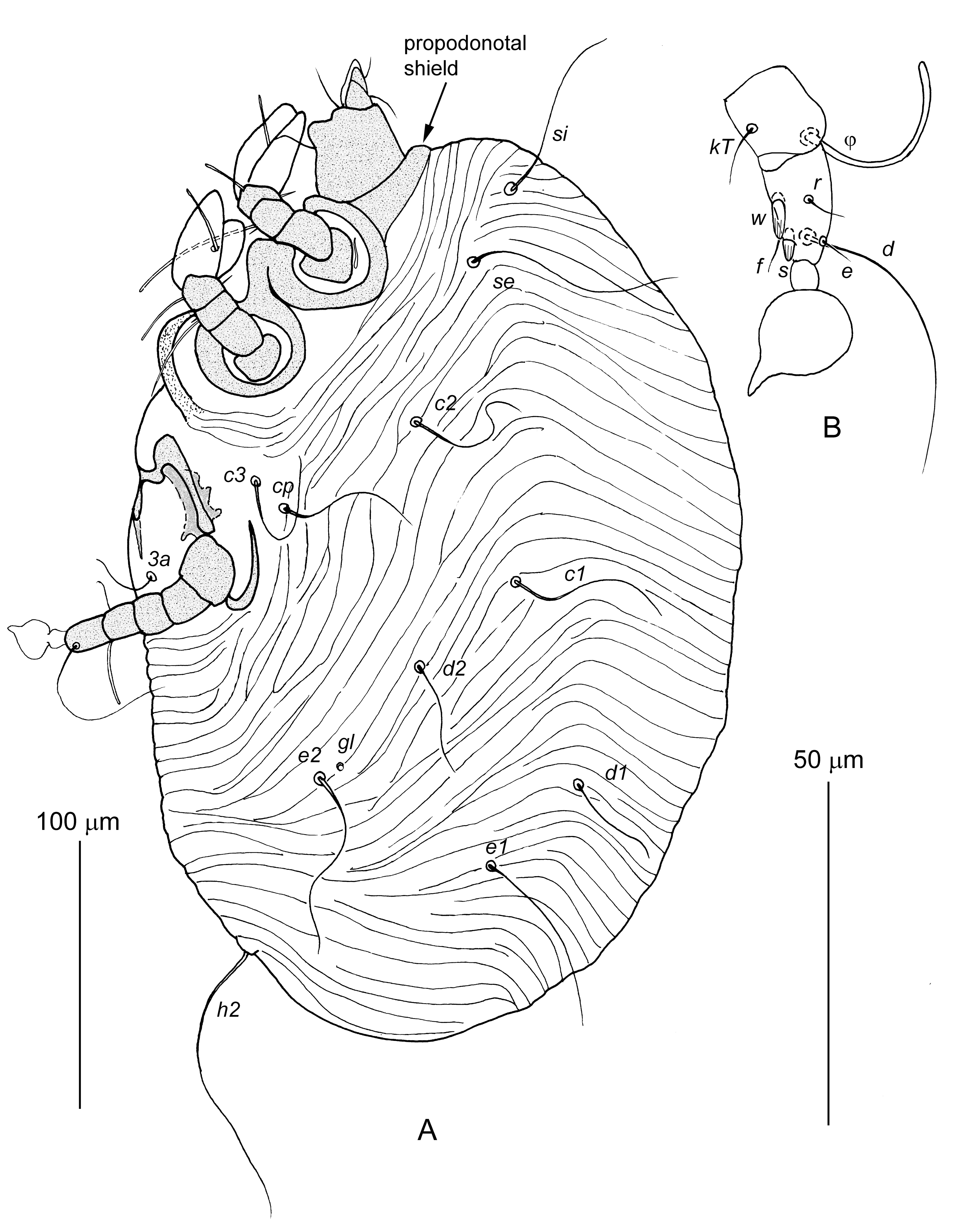
Male larva — (10 specimens, Figure 1) — Body 270–380 long and 165–255 wide. Gnathosoma. Gnathosoma having structure typical for Chirodiscidae with full complement of setae. Palps two-segmented, enclosed by membrane. Pseudorutellar membranes of subcapitulum weakly developed, dorsal lobes not developed. Palpal setae: dTi, dTil”, dTa, ω, ul’, and ul”; subcapitular setae: elcp and subc. Idiosoma. Idiosoma slightly flattened from lateral sides, completely covered by transverse striae. Propodonotal shield very short about 10 long, in shape of narrow transverse band. Hysteronotal shield absent. Hysteronotal gland openings gl distinct. Posterior margin of opisthosomal widely rounded. Laterocoxal setae scx absent. Anal opening situated ventrally. Lengths of setae: si 50–88, se 60–80, c1 55–63, c2 48–58, cp 45–50, c3 40–65, d1 33–45, d2 40–60, e1 50–65, e2 58–78, h2 150–210, 1a and 3a 15–20. Each pair of coxal apodemes Ia and IIa, fused in Y-shaped structures. Coxal apodemes IIIa fused medially in arch-like structure. Legs. Legs I and II with 5 segments: trochanter without setae, femur with seta vF, genu with setae cG and mG, tibia with seta gT and solenidion φ, tarsus with paired tarsal flaps and solenidion ω1. Legs III well developed, with full set of articulated segments: trochanter, femur, genu, tibia and tarsus with pretarsus. Ambulacral disc of pretarsus III with acute terminal protrusion. Leg III setation: genu: solenidion σ, tibia: seta kT and solenidion φ, tarsus: setae w, r, s, f, e, d. Setae sIII and wIII shaped as longitudinally striated spurs.
Male protonymph — (5 specimens, Figure 2A, B) — Body 310–390 long and 210–265 wide. Idiosoma. Propodonotal shield about 10 long. One pair of genital papillae, setae f2, h3, ps1, ps2, ps3, and g added on idiosoma. Setae f2 situated closer to bases of h2 than ps2. Lengths of setae: si 70–90, se 80–100, c1, c2, cp, c3, d1, d2, e1, and e2 70–100; f2, ps1, ps2, and ps3 10–20; h2 100–120; h3 80–100. Coxal apodemes IVa fused medially into arch-like structure. Legs. Legs IV with five articulated segments added. Pretarsus IV present. Setae d, w, and r of tarsus IV present, other segments of legs IV without setae. Seta wIV shaped as longitudinally striated spur.


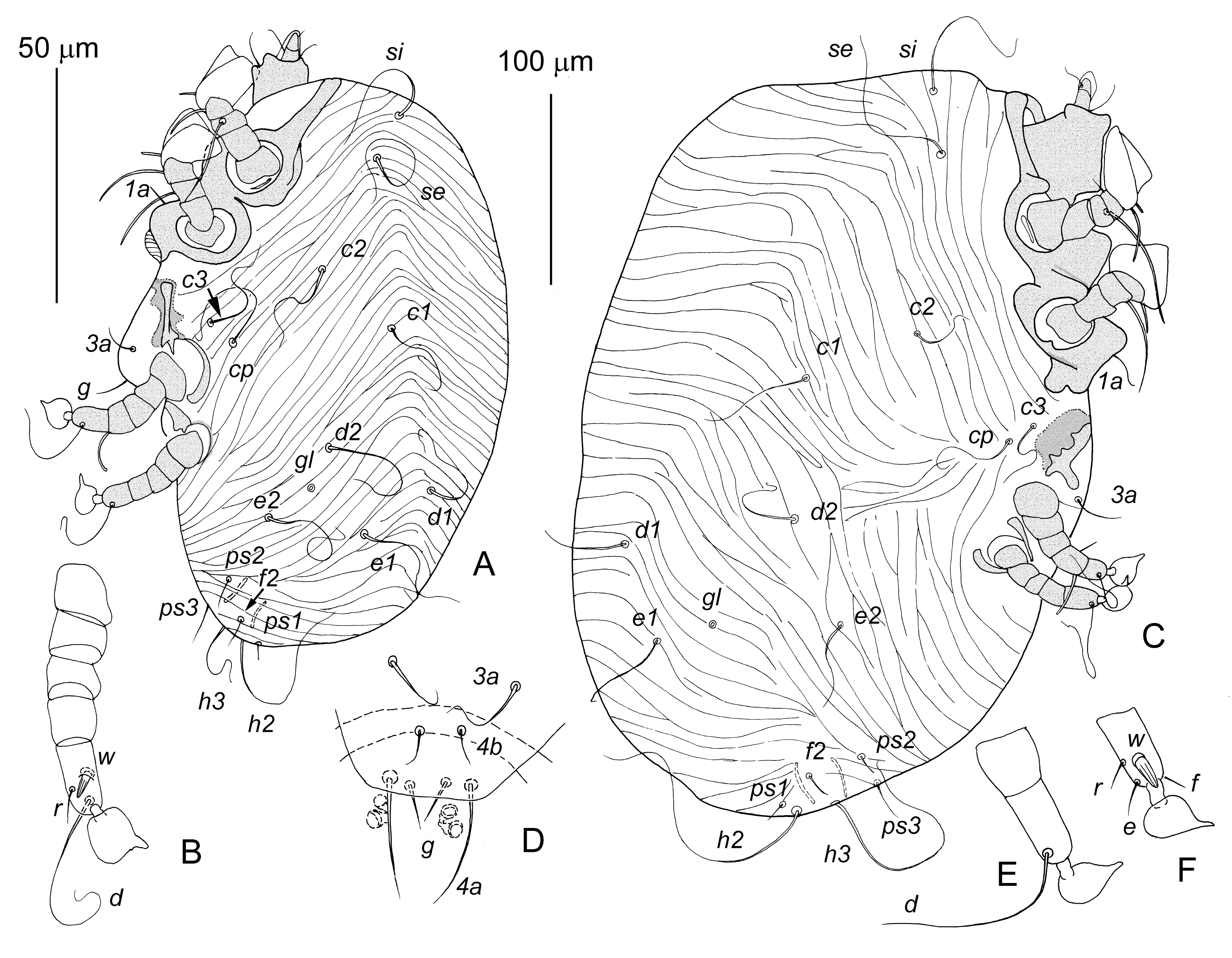









Male tritonymph — (10 specimens, Figure 2C–F) — Body 410–450 long and 250–300 wide. Idiosoma. Propodonotal shield 10–15 long. Second pair of genital papillae, coxal setae 4a and 4b added on idiosoma. Lengths of setae: si 78–90, se 80–100, c1 50–70, c2 70–90, cp 45–70, c3 60–65, d1 45–50, d2 70–90, e1 and e2 60–70, f2 18–25, h2 140–180, h3 80–100, ps1 5–10, ps2 23–28, ps3 38–53. Legs. Seta sRIII on trochanter III, seta kTIII on tibia III, and setae eIV and fIV on tarsus IV added.
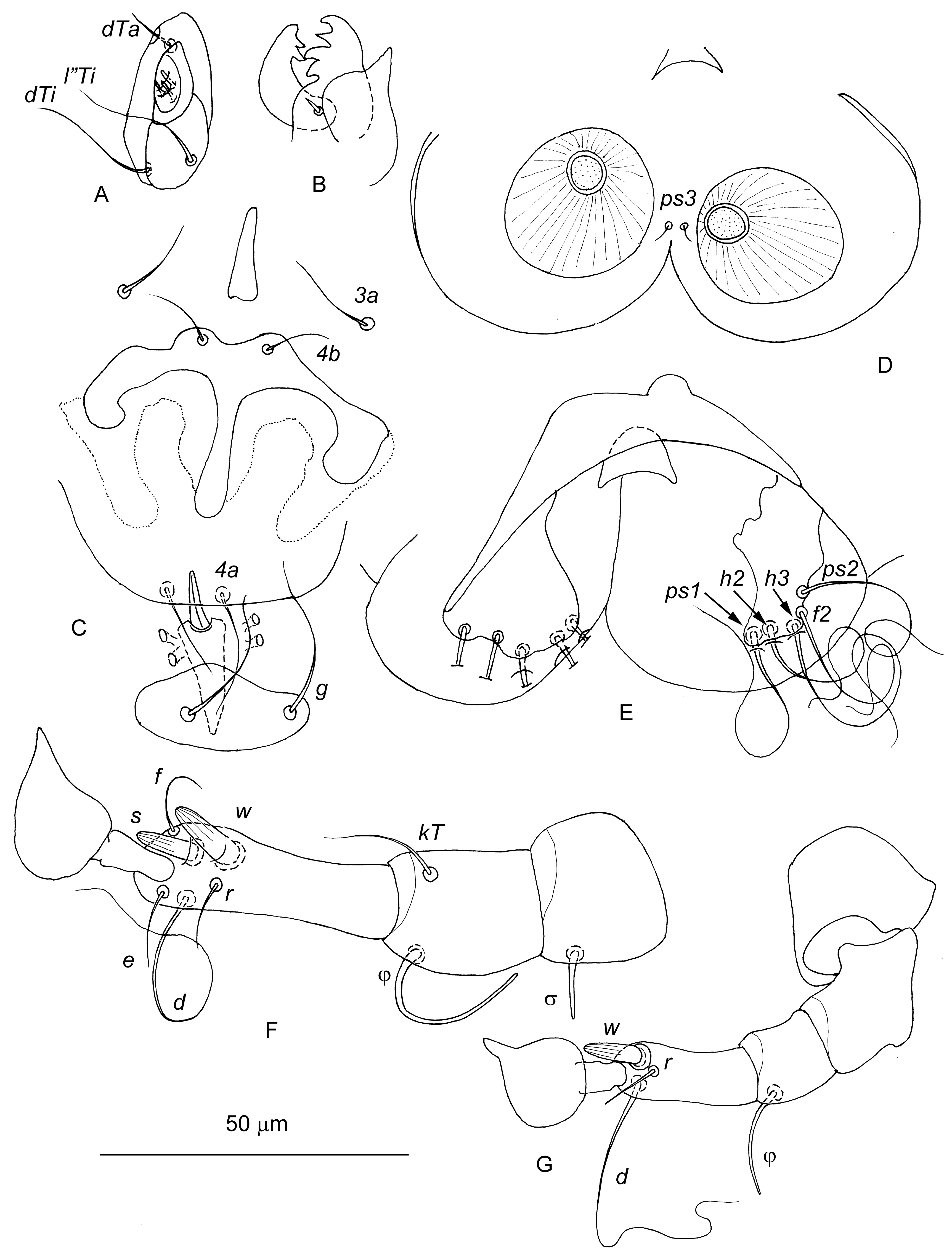
Male — (10 specimens, Figures 5A and 6) — Body 365–425 long, 205–265 wide. Idiosoma. Propodonotal shield 10–18 long. Idiosomal dorsum covered with numerous transverse striae. Hysteronotal shield shaped of transverse band, situated posterior to setae e1, 25–35 in length along midline. Setae ps3 situated between adanal suckers. Opisthosomal lobes widely rounded. Lengths of idiosomal setae: si 80–100, se 100–115, c1 78–93, c2 93–105, cp 70–120, c3 40–65, d1 40–60, d2 80–95, e1 – microsetae, e2 70–100, f2 80–85, h2 130–180, h3 130–170, ps1 110–120, and ps2 100–125. Aedeagus about 11 long. Adanal sclerites absent. Legs III 110–140 long, with setation as in male tritonymph. Legs IV 70–90 long, consisting of 4 segments, femur and genu fused. Solenidion σIII added on genu III. Tarsi IV bearing setae d, w, and r, tibia IV with solenidion φIV, other segments of legs IV without setae. Seta wIV shaped as longitudinally striated spur.
Female larva — (10 specimens, Figure 3) — Similar to male larva. Body 250–300 long and 160–205 wide. Idiosomal dorsum posterior to level of setal bases c2 and d1 smooth. Lengths of idiosomal setae: si 88–120, se 75–125, c1 28–38, c2 80–100, cp 80–120, c3 70–85, d1 2–3, d2 25–30, e1 110–125, e2 60–90, h2 180–240, 1a 10–20, and 3a 30–40. Setae e1 thickened and situated terminally. Live independently or attached to male.
Female protonymph — (10 specimens, Figure 4A, B) — Body 255–265 long and 190–210 wide, slightly elongated sack-like. Coxal fields I smooth; coxal apodemes II–IV absent. Setae c3, e1, e2, d1, d2, f2, 3a, g, ps1, ps2, and ps3 absent. Setae h3 added. Lengths of setae: si 88–93, se 100–110, c1 10–15, c2 5–8, cp 30–35, 1a about 10, h2 and h3 28–35. Posterior end of opisthosoma between levels of setae e1 and h2 smooth, anterior margin of this area flanked with a pair of large bow-shaped opisthosomal sclerites and bears pair of rounded tubercles corresponding to male adanal suckers. Legs I strongly shortened compared to female larva, their tibia and tarsus clearly separated. Setation of legs I as in female larva, but all setae shorter. Legs II–IV absent. First pair of genital papillae added.
Female tritonymph — (10 specimens, Figure 4C) — Similar to female protonymph. Body 335–350 long and 245–305 wide. Idiosoma. Propodonotal shield 10–15 long. Second pair of genital papillae added. Idiosomal chaetome as in female protonymph. Lengths of idiosomal setae: si 80–135, se 125–155, c1 and c2 5–8, cp 25–50, h2 and h3 20–35, 1a about 10. Legs. Legs I as in female protonymph. Legs II strongly reduced, several times shorter and thinner than legs I, consisting of 3 articulated segments, apical segment with 3 short setae. Legs III and IV primordial, shaped as small conical tubercles not split into segments, each bearing 1 apical microseta.
Female — (10 specimens, Figures 5B and 7) — Similar to male tritonymph except oviporus between coxal fields III. Body 440–490 long, 275–330 wide. Idiosoma. Propodonotal shield about 10 long. Full set of idiosomal setae occurring in adult chirodiscids excluding setae h1 (as in male). Coxal apodemes I–III as in male tritonymph. Lengths of setae: si 100–125, se 100–120, c1 75–85, c2 80–105, cp 83–105, c3 75–85, d1 58–80, d2 85–95, e1 95–130, e2 95–125, f2 19–25, h2 135–170, h3 140–160, ps1 10–23, ps2 25–38, and ps3 50–70. Legs. Legs I and II as in male. Legs III and IV fully developed as in male tritonymph; setation of legs III and tibia IV as in male, tarsus IV with 5 setae d, e, f, r, and w. Seta wIV shaped as longitudinally striated spur. Ovoviviparous.
I thank Dr. A.V. Abramov (ZISP), who collected the host specimen. This research was supported by the Russian Foundation for Basic Research (Grant No. 16-04-00085a).
Editorial note: This contribution was reviewed after the untimely death of its author; hence we leave it without Discussion, as originally submitted. Helpful suggestions of an anonymous reviewer are greatly appreciated and, apart from the one related to the Discussion, accepted. Generous assistance of Dr. S.V. Mironov (ZISP) in leading the manuscript through the pre-publication process is gratefully acknowledged.
Bochkov A.V. 2010. A review of mammal-associated Psoroptidia (Acariformes: Astigmata). Acarina, 18: 99-260.
Bochkov A.V., Mbalitini S.G., Verheyen E. 2016. Review of the fur-mite genus Soricilichus Fain, 1970 (Acariformes: Chirodiscidae). symbionts of the African shrews of the subfamily Crocidurinae (Soricomorpha: Soricidae). Zootaxa, 4072: 235-253. doi:10.11646/zootaxa.4072.2.6 ![]()
Dubinina E.V. 1964. Development cycle of the mites of the genus Histiophorus (Sarcoptiformes : Listrophoridae). Zool. Zh., 43: 534-548. (in Russian)
Evans G.O. 1992. Principles of Acarology. Wallingford, UK: C.A.B International, pp. 563.
Fain A. 1970. Diagnoses de nouveaux lobalgidés et listrophoridés (Acarina: Sarcoptiformes). Rev. Zool. Bot. Afr., 81: 271-300.
Fain A. 1971. Les listrophorides en Afrique au sud du Sahara (Acarina: Sarcoptiformes). II. Familles Listrophoridae et Chirodiscidae. Acta Zool. Pathol. Antv., 54: 1-231.
Fain A. 1981. Notes sur des acariens des genres Soricilichus Fain et Lutrilichus Fain (Acari: Chirodiscidae). Bull. Ann. Soc. R. Ent. Belg., 117: 41-44.
Fain A., Lukoschus F.S., Kock N.J.J., Glulow F.V. 1974. A key to the genus Lutrilichus Fain and description of a new species from the ermine, Mustela erminea, in Canada (Acarina: Sarcoptiformes). Can. J. Zool., 74: 941-944. doi:10.1139/z74-125 ![]()
Grandjean F. 1939. La chaetotaxie des pattes chez les Acaridiae. Bull. Soc. Zool. Fr., 64: 50-60.
Griffiths D.A., Atyeo W.T., Norton R.A., Lynch C.A. 1990. The idiosomal chaetotaxy of astigmatid mites. J. Zool. (London), 220: 1-32. doi:10.1111/j.1469-7998.1990.tb04291.x ![]()
Norton R. 1998. Morphological evidence for the evolutionary origin of Astigmata (Acari: Acariformes). Exp. Appl. Acarol., 22: 559-594. doi:10.1023/A:1006135509248 ![]()



2017-12-13
Date accepted:
2018-06-15
Date published:
2018-09-10
Edited by:
Kreiter, Serge

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Bochkov, Andre V.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)