




Paraquanothrus n. gen. from freshwater rock pools in the USA, with new diagnoses of Aquanothrus, Aquanothrinae, and Ameronothridae (Acari, Oribatida)
Norton, Roy A.1 and Franklin, Elizabeth2
1✉ State University of New York, College of Environmental Science and Forestry, Syracuse, New York, 13210, U.S.A.
2National Institute for Amazonian Research, Biodiversity Coordination, CEP 69067-375 Manaus, Amazonas, Brazil
2018 - Volume: 58 Issue: 3 pages: 557-627
https://doi.org/10.24349/acarologia/20184258ZooBank LSID: A9FDC4A9-B70D-4965-9F7E-94813BB2929D
Keywords
Abstract
Oribatid mites form a dominant component of the soil fauna, being particularly abundant and diverse in complex organic horizons of moist forest floors, where they feed primarily on decaying plant remains and fungi. But several important higher taxa are rarely represented in such habitats; among them are four nominal superfamilies in the hyporder Brachypylina—Licneremaeoidea, Cymbaeremaeoidea, Ameronothroidea and Limnozetoidea—that have adapted to rather different conditions (Weigmann 2006; Norton and Behan-Pelletier 2009).
Xeric soils, and microhabitats that undergo extreme hydric variation regardless of general climate (e.g. arboreal and rock surfaces, exposed mosses and lichens), are typical homes for members of the first two superfamilies, which overall could be considered ‘dry-adapted’ or ‘dry-tolerant’ (Norton and Behan-Pelletier 2009). By contrast, many members of the third and fourth superfamilies are ostensibly ‘wet-adapted’ or ‘wet-tolerant’. Limnozetoid species are aquatic or semiaquatic in freshwater habitats, and many ameronothroid species are marine-intertidal (Schulte et al. 1975; Behan-Pelletier and Eamer 2007; Pfingstl 2017).
But differences in life-style and niche-adaptation among members of these groups are not always as great as such generalities suggest. Oribatid mites in exposed terrestrial microhabitats—e.g. Scutovertex and Scapheremaeus species living in arboreal or epilithic mosses and lichens—may be inactive during dry periods (Travé 1963a; Walter 1999; Smrž 2002; Colloff 2010). Conceptually, this differs little from ‘aquatic’ species in the ameronothroid genera Aquanothrus and Chudalupia, which become immobile when their temporary rock pool habitat no longer contains water (Engelbrecht 1975; Wallwork 1981). At least one species of Scapheremaeus, S. baylyi Colloff, 2010, appears to have a similar lifestyle in rock pools (Colloff 2010). Even the habitats themselves are not necessarily discrete: for example, another species of Scapheremaeus seems to live only in frequently-dried mosses that grow at the edge of exposed, ephemeral rock pools (Norton et al. 2010).
Our main purpose is to describe and discuss two new species inhabiting such rock pools (= lithotelmata, gnammas, pans). These are small, temporary lentic environments formed by the accumulation of rainwater in shallow depressions—typically solution or weathering pits—on rock surfaces. They provide habitat for aquatic arthropods and other small invertebrates capable of surviving extended periods of desiccating conditions (Bayly 1997, Jocqué et al. 2010 and cited references). For reasons discussed below, we propose Paraquanothrus n. gen. to include these new species and consider it the sister-genus of Aquanothrus, the type species of which—A. montanus Engelbrecht, 1975—inhabits rock pools in South Africa (Engelbrecht 1975; Jocqué et al. 2006; Vanschoenwinkel et al. 2009).
The two new species are endemic to widely separated locations in the USA that present very different environmental conditions. Paraquanothrus grahami n. sp. is restricted to the arid Colorado Plateau, where it inhabits shallow depressions on exposed sandstone. Paraquanothrus spooneri n. sp. is known only from depressions formed in exposed granite outcrops in the warm-temperate state of Georgia. As with A. montanus, adults and juveniles of both species are active only when immersed in water; they burrow into sediment as it dries and remain immobile until water returns. Mites of both genera have been called ‘cryptobiotic’ in the literature (Vanshoenwinkel et al. 2009; Colloff 2010) but they cannot endure desiccation: they survive by preventing it.
Secondary objectives are to: (1) improve knowledge of Aquanothrus montanus by adding ontogenetic data, correcting some errors in the original description, critiquing the supposed high variability of the species, and proposing a new generic diagnosis; (2) discuss the systematic position of Paraquanothrus and Aquanothrus by reviewing and critiquing past concepts of Ameronothridae and recasting Aquanothridae as a subfamily of Ameronothridae; and (3) evaluate morphological support for molecular studies that have brought the monophyly of Ameronothroidea, Cymbaeremaeoidea and Licneremaeoidea into question.
Acquisition and deposition of specimens — Active adults and juveniles of the two new species were collected directly from rain-filled rock pools by eyedropper or similar device. Inactive specimens were obtained from dry rock-pools by gently collecting sediment. In most cases, this material was rehydrated with rainwater or spring water in the laboratory, then the revived mites were observed alive under a stereomicroscope and manipulated with a fine brush or collected by pipette. Collection data are given below. Numerous preserved adults and juveniles identified as Aquanothrus montanus were studied, including paratype specimens from each of the three South African sites indicated in the original description (Engelbrecht 1975), as well as adults and juveniles from Botswana (Thamaga), a population studied by Jocqué et al. (2006). Preserved adults and nymphs of Chudalupia meridionalis Wallwork, 1981 from the type locality in Western Australia were studied. Specimens of other species discussed below are from the general collection of the first author. Type repositories include the mite collections of: the United States National Museum (USNM), the Smithsonian Institution (Washington, D.C.), currently with the US Department of Agriculture (Beltsville, Maryland); the Canadian National Collection (CNC), Agriculture and Agri-foods Canada, Ottawa; and the Instituto Nacional de Pesquisas da Amazônia (INPA), Manaus, Brazil.
Specimen preparation and documentation — Most observations and data are from specimens temporarily mounted in cavity slides in a medium of lactic acid diluted with water (2:1; Grandjean 1949b). Dissected mouthparts and legs were permanently mounted in Hoyer's medium for observation with oil-immersion lenses. Compound microscopy employed bright-field, polarized, and Nomarski (DIC) illumination; polarized light was particularly employed to distinguish setae (birefringent) from solenidia (isotropic) when correcting literature statements. Line drawings were made with the aid of a drawing tube. Light micrographs were obtained, usually as image stacks, with a Nikon CoolPix 995 or AmScope MU800 digital camera mounted on a Nikon Ellipse 800 compound or SMZU stereomicroscope. Image stacks were combined using the Helicon Focus Pro (v. 5.0) suite; the stacks varied widely in number of individual images, usually only several for highly magnified (1000 x) images and 15-30 for lower magnifications. As needed, images were adjusted with Adobe Photoshop (CS3) for contrast and color balance. Frames for the supplemental video file were acquired with the AmScope camera and stereomicroscope and processed with OpenShot video-editing software. Body length was measured in dorsoventral aspect, from the tip of the rostrum to the posterior edge of the hysterosoma. Unless otherwise noted, width refers to the maximum width in dorsal aspect (found on the hysterosoma). Measurements of specific structures or distances are meant to be representative of an average-sized individual, though a range is often given. Setal measurements include only the freely emergent part (omitting alveolus or apobasic portion).
Terminology and nomenclature — Morphological terminology is mostly that of F. Grandjean (see Travé and Vachon 1975 for references and Travé et al. 1996 or Norton and Behan-Pelletier 2009 for overview). Terms are translated from French (Hammen 1980) but in most instances Grandjean’s original abbreviations and figure notations are retained. The notations used for epimeral setae are slightly modified (see Remark 15), and a few terms and abbreviations are from Schubart (1975). Setal formulas are given as counts per segment for appendages (from femur to tarsus) and per podosomal segment (I-IV) for epimeres. Paired structures are described in the singular unless noted. Unless otherwise indicated, parentheses around leg setal notations denote the two members of a pseudosymmetrical pair on a given leg segment, rather than a true bilateral pair. Taxonomic context is that of Norton and Behan-Pelletier (2009) and Schatz et al. (2011). Author and date for species-group taxa are given at the first use of the name; those of supraspecific taxa can be found in Subías (2004).
Supplementary material — this article has a video file associated with it.
ZOOBANK: 8A9E6335-F837-4092-B39E-BBB373465E4A ![]()
Type species — Paraquanothrus grahami n. sp.
Diagnosis — With traits of Aquanothrinae (see below). Adult. Invaginated portion of apobasic setal insertions in form of small cup. Bothridial opening flush with surface; bothridial seta rod-like, distally acute; seta ex variable within populations, never longer than le, often minute or absent. Dorsosejugal scissure complete, anterior margin of notogaster evenly arched in dorsal view. Lenticulus small, width less than one-third mutual distance of setal pair c1. Notogaster with 15 or 13 pairs of setae (p1, p2 present or absent; c1-3 always present). Ovipositor without coronal setae. Usually two pairs of anal setae, inserted in posterior half of plate. Exposed part of preanal organ (preanal plate) clearly wider than long, about same width as hidden vertical wall. Palp with large basal tubercle bearing setae cm, acm and solenidion ω; setal pair (ul) eupathidial, acm and su normal. Postpalpal seta ep spiniform. Leg pretarsi monodactylous. Trochanters III, IV with basal articulating stalk attached subterminally. Porose organs on all femora invaginated (tracheae or saccules), those of trochanters III-IV invaginated or surficial. Famulus present, small; seta ft″ of tarsus I close to solenidion ω1 in same or separate alveolus; solenidion ω of tarsus II coupled with seta ft′ in same alveolus, ω2 absent; proral setae eupathidial on tarsus I; iteral setae of tarsus IV absent, those of I-III present or absent; tarsi without or with at most one primiventral seta. Seta d absent from all genua. Trochanters I, II, IV glabrous. Juveniles. With four pairs of distinct or vague patches of leathery cuticle: in lateral, aggenital and posteroventral regions, and associated with opisthonotal gland opening. Line of dehiscence (δ) passing above cupule ip in both larva and nymphs. Protonymphal leg IV setal formula 0-0-0-0-6 (pv″ absent). Egg with external layer of dense microgranules, ca. 1 μm diameter.
Adult
Facies (Figs 1, 2, 9) — Medium to large mites (ca. 650-900 μm long). Narrowly ovate in dorsal view; notogastral length about 3.2-3.5 times that of prodorsum; maximum width about 1.4-1.7 that of prodorsum. Somewhat ‘blunt-faced' in lateral view, with relatively long hysterosoma (2.5 or more times length of proterosoma). Legs proportionally rather small, thin. Genders with approximately equal frequency; no obvious sexual dimorphism except in genitalia.
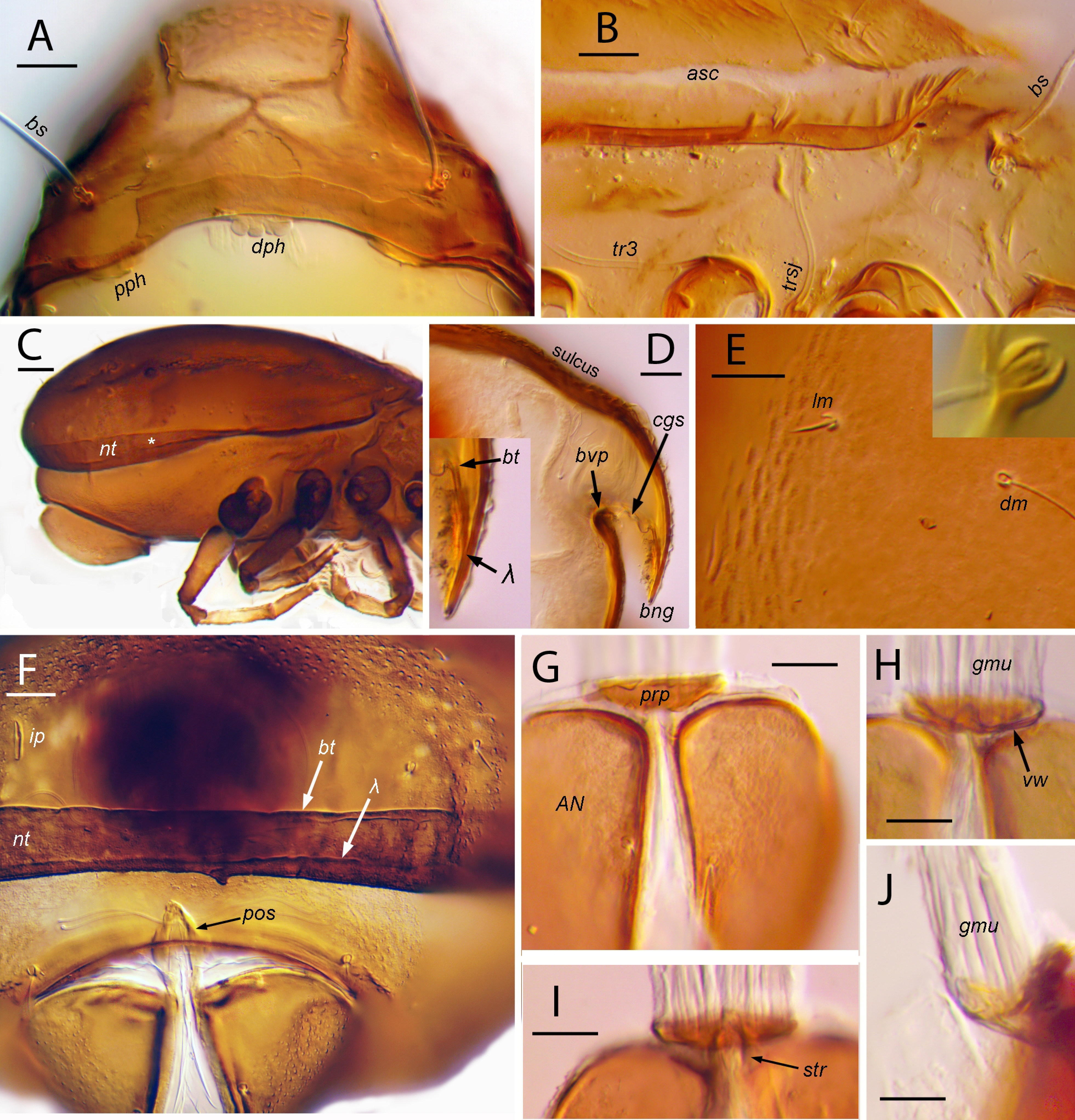
Cuticle — Weakly (easily deformed) to modestly sclerotized, smooth or with various folds or wrinkles. Cerotegument uniformly covering body and legs (except as noted below); without conspicuous projecting excrescences, but with complex structure of dense, distally expanded, minute (less than 1 μm diameter) papillae, giving cuticle ‘sanded' appearance in transmitted light (Figs 4A, 4B left edge); papillae closely adjacent, forming micro-chambered layer above procuticle (Fig. 3B, D, E). Cerotegument strongly pigmented in mature adults: nearly black in life, medium to light brown after long preservation, except pale notogastral lenticulus. Underlying procuticle pale (Figs 4B, 16C), with only weak, inconspicuous porosity (Fig. 3E) except within porose areas (Fig. 12I). Setae of body mostly apobasic, with insertion at bottom of internally-projecting, barrel-like cup (Figs 4B, 10E); some proximal leg setae apobasic or not. Cuticle without crust of adhering epibionts: clean or with small scattered debris or occasional diatoms.
Prodorsum — Relatively short, occupying only ca. 20% of body length in dorsal aspect; outline roughly triangular in dorsal aspect, strongly convex in lateral view; lamellar seta (le) inserted approximately where prodorsal curvature becomes nearly vertical, appearing close to rostral seta (ro) in foreshortened dorsal aspect (Figs 1A, C, 9A, C); interlamellar (in) and exobothridial (ex) setae regressive and variable (small, minute or absent). Bothridium narrow, tubular throughout, ca. 7-8 μm wide, weakly S-shaped, opening flush with surface and inconspicuous under small cuticular fold; bothridial seta (bs) tapered only near tip, weakly barbed (Figs 1D, 4A, 9D). With three small, inconspicuous apodemes extending from posterior margin (Fig. 10A) — medial dorsophragma (dph) and pair of rudimentary pleurophragmata (pph).
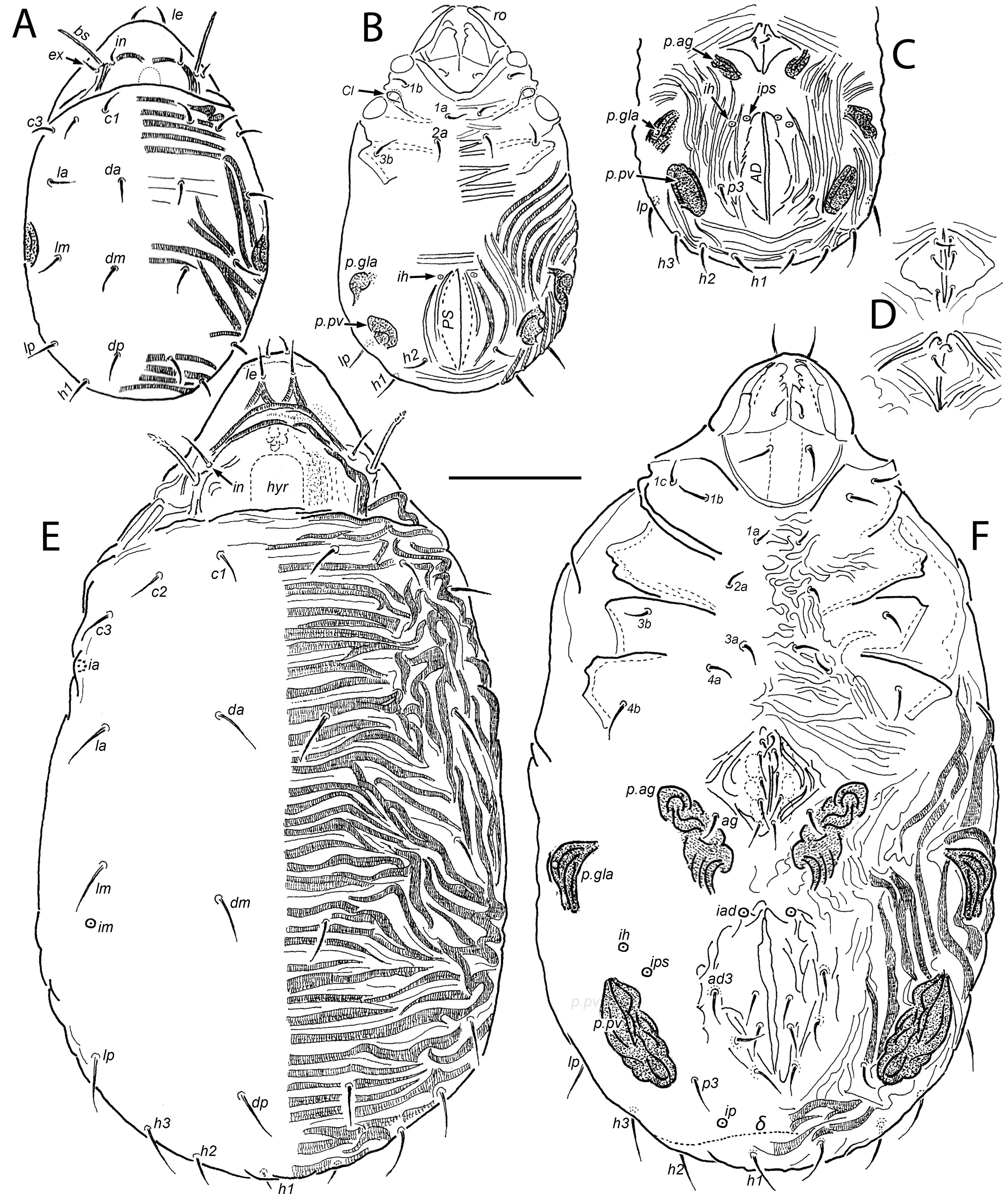
Notogaster — Outline in dorsal view simple, elliptical to slightly ovate, 1.3-1.5 times longer than maximum width; evenly convex anteriorly, not projecting forward or merged with prodorsum (Figs 1A, B, 9A, C). With well or poorly defined central dome and rim; border with or without posterolateral tectum. Cuticle immediately around opisthonotal gland opening often darkened; gland rather flat, purse-like. Lenticulus conspicuous, sharply circumscribed, convex; relatively small, width less than one-third mutual distance of setal pair c1; cuticle hyaline, birefringent in polarized light; with paired spots of underlying red pigment in life (Fig. 16A). With 15 or 13 pairs of setae (p1, p2 present or absent; c1-3 always present); setae uniformly small, thin, smooth.
Podolateral and coxisternal regions — Distinct pedotecta absent. Without discrete discidium, but oblique fold (of; Figs 1B, C, 9B, C) present between acetabula III, IV (‘discidial ridge' of authors). Tracheal system normal: that of acetabulum I and sejugal furrow bifurcated, that of acetabulum III single (Fig. 4E); basal region of trachea 3 usually swollen, lined with dark cerotegument. Coxisternum with distinct, transverse sejugal furrow and similar furrow between epimeres I and II (Fig. 2B). Crescent-shaped mentotectum of epimere I very narrow. Apodemes variously developed (Fig. 4C, D): ap1 continuous between acetabula but broadly U-shaped, laterally high with thickened edge, medially low and inconspicuous. Other apodemes formed as paired demiapodemes, effacing prior to midline and perforated (troué) lateroventrally (i.e., attached to acetabulum laterally but to respective epimere only at medial end); ap2 large, bladelike, with thick edge, rising to peak near medial end just anterior to insertion of seta 2a; apsj slightly shorter but much less developed, maximum height in lateral region, obliquely effacing before reaching level of 2a; ap3 strut-like, descending obliquely from acetabulum III to efface no more than halfway from acetabulum to seta 4a. No apodeme 4 developed. Epimeral setation 3-1-2-2; position of seta 4a slightly variable, sometimes inserted closely posterolateral to 3a (giving apparent setation of 3-1-3-1).
Anogenital region — Anal aperture ca. 1.3-1.4 times longer than genital aperture; both nearly as wide as long and separated by slightly less than length of genital aperture; genital aperture of male slightly smaller and more distant from anal aperture than in female. Genital plate with small internal nodule at posteromedial corner, receiving muscle tendon from preanal organ; six pairs of genital setae aligned near medial margin, with posterior seta on posteromedial corner, distant from others. Aggenital seta present; with three pairs of adanal and usually two pairs of anal setae (variation noted below); no ventral neotrichy. Preanal organ (Figs 4G, 10G-J) with preanal plate (prp), wider than long, mostly visible externally in anterior fold of anal vestibule; vertical wall (vw), serving as origin of genital muscles (gmu), joining posterior edge of prp at right angle; supporting strut (str) rising vertically from anteromedial corner of each anal plate, pair well separated for most of their length, attaching near top of vw. Posteromedial corner of each anal plate with similar supporting strut rising dorsally, merging to form narrow, simple, ‘hairpin-like' postanal strut (pos), serving as insertion for postanal suspensor muscles (Figs 4F, 10F). Apparent medial edge of anal plate sharply flexed dorsad at right angle, creating narrow vertical wall. Anterior anal seta approximately aligned with seta ad3, well posterior to level of lyrifissure iad; iad slit-like, aligned with and closely adjacent to margin of anal aperture. Genital papillae of uniform size, about three times longer than wide in cross-section (Fig. 4C, insert). Ovipositor (Fig. 6B) without coronal setae; distal setae (τ1 on each paired lobe and pair ψ1 on unpaired ventral lobe) distinctly shorter than respective lobe, not extending beyond its tip.Spermatopositor of normal short form, with five or six pairs of setae (ψ2 present or absent). Eggs with external layer of dense microgranules, ca. 1 μm diameter (Fig. 6L).
Subcapitulum — Length (including rutellum) 1.2-1.3 times breadth; labiogenal articulation complete or laterally slightly incomplete (Figs 5C, 11C). Gena short, subtriangular, only ca. 1/4 to 1/3 as long medially as laterally; procuticle porose. Rutellum short, with large thumb-like dorsolateral process, typical dentition, and well-formed rutellar brush of strong, almost spiniform cilia; without rutellar oncophysis; atelobasic, with large medial lobe, pair not overlapping at midline (unless distorted in preparation). Mentum porose anteriorly; hypostomal (h) and genal setae (a, m) apobasic; h inserted anteriorly on mentum, pair close together with mutual distance about same as genal setae a. Adoral setae with normal insertion; strongly curved dorsad, tapered but not finely attenuate; with short, inconspicuous barbs. Postpalpal seta (ep) spiniform. Labrum broad throughout, rounded distally, without cilia or denticles; medial longitudinal crest distinct above narrow, distally tapered, embedded labral sclerite. Opening to paired subcapitular (infracapitular) gland distinct. Capitular apodeme with typical bilobed form, but very thin and inconspicuous; obtusely angled in cross-section (Fig. 12E).
Palp (Figs 5D, 6C, 11D) — With typical five freely articulated segments; distal four segments seem without cerotegument, with clearly porose procuticle. Setation 0-2-1-3-9, plus tarsal solenidion ω; setae not apobasic. Femoral seta sup in mid-lateral position. Tarsus with dorsal apophysis in proximal third, abruptly narrowing to cylindrical, slightly bowed distal region having about half height of proximal third in lateral view. All tarsal setae relatively short: cm and acm at top of proximal tubercle, pair (lt) more distal, at tubercle base; ul″ dorsal, near midpoint of cylindrical distal region of tarsus, well removed from subterminal ul′; vt′ near ventral midline, vt″ on abaxial face, su terminal. Only ultimal pair (ul) eupathidial. Solenidion ω on proximal tubercle, abaxial of and about equidistant from cm and acm; not coupled to either (no ‘double-horn'). Tarsal lyrifissure present but inconspicuous (not illustrated).
Chelicera — Form typical of family (Figs 5E, 11E); each digit with four strong teeth (including terminal teeth and lateral subterminal tooth of fixed digit). Without cerotegument; procuticle strongly porose distal to attachment of cheliceral frame. With long, internally looping, serpentine trachea ending in gradually expanding, narrow bulb (Fig. 6N). Setae pigmented except near base and densely barbed: cha larger, inserted dorsally, slightly on adaxial face and slightly distal to mid-length; chb about half size of cha, inserted in mid-lateral position on abaxial face, just posterior to fixed digit. Trägårdh’s organ (Tg) narrowly tapered, with distinct, often noticeably porose sclerite (Fig. 6M).
Legs — Relatively short: leg IV longest, about 0.5-0.7 times body length, relatively thin (Figs 6A, 12A). Trochanters III, IV short, distally widening in lateral aspect; less than 50% longer than maximal height (in distal half) and height ca. 50% greater than respective femur. Right-angled proximal stalk of trochanters III, IV subterminal, with proximal end of segment extending to cover insertion into acetabulum; most striking on trochanter III, where flat adaxial and convex abaxial face resembles asymmetrical mushroom in cross-section (Fig. 12D). Femora I, II gradually broadening from narrow base to maximum width at mid-length; femora III, IV subrectangular with abrupt proximal constriction, forming short stalk; femur IV ca. 3 times as long as broad. Tibiae III, IV conspicuously longer than I, II: tibia IV ca. 4-5 times as long as broad. Tarsi I, II short (ca. 1.7 times length of claw), bulging in proximal half, strongly tapering in distal half to form short stalk; tarsi III, IV longer, without proximal bulge, distal half narrowing gradually to base of claw. Midventral cuticle of all tarsi with guide channel for claw-depressor tendon, extending short distance proximally as free tube (Fig. 6F). With internalized porose organ (saccule or trachea) opening adaxially on each femur; trochanters III, IV with trachea or porose area; tibiae and tarsi without porose organ. Pretarsi monodactylous; proximal half of claw with paired dorsolateral row of minute denticles (Fig. 5B). Leg setation partly regressive: trochanters I, II and IV glabrous; seta d absent from all tibiae and from genua I-III; primilateral pair (pl) absent from tarsus I; primiventral pair (pv) absent from tarsi II-III, pv′ absent from tarsus I, pv″ absent from tarsus IV; iteral setae (it) absent from tarsus IV, present or absent on I-III. Setae acute to attenuate, of moderate length, many as long as or longer than segment width. Seta d of femora I, II inserted distinctly on abaxial face, nearly aligned with seta l′. Unguinal setae (u) weakly s-shaped; basally thick and coarsely barbed, distally finely attenuate (often broken), curving parallel to claw. Proral setae (p) minute, eupathidial on tarsus I, reaching only to base of claw; pair normally formed, barbed, on other tarsi. Subunguinal seta s not eupathidial. Tectal (tc) setae (and iterals if present) long, reaching past claw (except on tarsus IV of P. grahami), weakly curled ventrad at slightly thickened tip (Fig. 6I); fastigial setae (ft) shorter, often only about half as long as (tc), and not distally curled. Disjunction of tectal pair (and of fastigials, iterals when pair present): ″ on tarsus I, ″ or absent on II, ′ or absent on III, IV. Famulus e of tarsus I small to minute, with thread-like internal ‘root’ (Fig. 6E). Seta v′ of tibiae I, II thickened, stiff, conspicuously barbed. Solenidia relatively short, thin, mostly baculiform, isodiametric except longest solenidion (φ1 of tibia I) very slightly tapered (ceratiform); tarsus II lacking second solenidion. Solenidial counts: genua 1-1-1-0; tibiae 2-1-1-1; tarsi 2-1-0-0. Solenidion ω of tarsus II coupled with seta ft′ within same alveolus.
Juveniles
Cuticle — Cerotegument thin, uniform throughout (exceptions as in adult); structured as in adult or with papillae coalesced, chambers opening to surface through pores (Fig. 3G); dark in life (Fig. 8C) except prodorsal aspis with hyaline region (hyr) posteromedially (Fig. 8B). Mostly pale in teneral period or after long preservation, with dark color persisting locally, or not, according to species. Procuticle porosity difficult to discern beneath chambered cerotegument, but limited in distribution: present in trapezoidal interbothridial region of prodorsum (except for hyaline region), in parts of epimeral plates, in leathery patches of hysterosoma (at least in P. spooneri) and in porose areas of leg femora (see below). Setae of various forms, but none apobasic. Most simple setae of body and legs (referred to below as ‘collared’) with base partly enclosed in darker, tubular collar (Figs 8F-G, 12H).
Prodorsum — With weakly defined creases in distal half; most of sub-trapezoidal basal half smooth, with underlying porosity. Aspis distally with broad lobe bearing rostral setae (hidden in dorsoventral view). Setae as in adult, including setiform bothridial seta and regressed bothridium, except non-bothridial setae may be collared. Setal pairs in and ex consistently present.
Gastronotic region — Dorsally convex in cross-section, without central dome or lateral rim but may be somewhat depressed at insertions of major dorsoventral muscles (artificially exaggerated in Fig. 3F). Gastronotic plication generally transverse (longitudinal in mid-lateral region), finely to moderately spaced; without porose dorsal sclerites (Fig. 17A, C), but with four paired lateral and ventral patches of leathery-appearing cuticle (Fig. 17B, D), distinguished by somewhat less-defined or absent plication; patches with or without strong, persistent coloration in cerotegument. Patches include: elongated lateral patch (p.lat) in region ventral to setae la, lm; smaller irregular or rounded periglandular patch (p.gl) associated with opening of opisthonotal gland (gla); variously shaped posteroventral patch (p.pv); and aggenital patch (p.ag) lateral to seta ag. Gastronotic setation of larva uni- or bideficient (h3 present or absent, according to species), that of nymphs uni- or trideficient (3 or 1 pairs of setae in p-row); setae homogeneous, short as in adult. Cupule ia posterior or posterolateral to seta c3, at level between c3 and anterior end of patch p.lat; cupule im aligned with dorsolateral setal row, ca. 1/4 to 1/3 distance from lm to lp; ip posterodorsal to paraproctal valves. Ontogeny of other cupules normal: ih, ips, iad just lateral to anterior end of paraprocts in larva, protonymph and deutonymph, respectively, with ih and ips displaced laterally in later nymphs. Line of dehiscence (δ) U-shaped, running around hysterosoma posteriorly (Figs 7F, 8H, 13F), resulting in one-piece exuvium (Fig. 8I); δ passing dorsal to gla and ip in both larva and nymphs, then following dorsal margin of patch p.lat (Fig. 12F) and effacing at level midway between setae la and lm; distance between δ and patch p.gl at least as great as diameter of patch.
Coxisternum — With weakly sclerotized epimeral plates, broadly divided medially by plicate cuticle (Figs 7B, 13F); plicae mostly short, irregular, except mostly transverse posteriorly; medial margin of plates imprecise. Epimeral groove 2 long, deep, narrow; procuticle of anterior groove wall porose, porosity extending anteriorly part-way across first epimeral plate; sejugal groove similar but porose on both walls, with porosity extended part way onto epimeral plates II and III; epimeral groove 3 shorter, with porosity only on posterior wall, extending part way across epimeral plate IV. Insertions of legs I-III (not IV) with rudimentary acetabulum, invaginated wall porose. Condyle K (articulating with subcapitulum) relatively large, tongue-like, extending anteriorly from dorsal region of epimere I (above leg insertion); podocephalic canal running along dorsal edge of epimere, across condyle and onto dorsal surface of subcapitulum in typical manner. Larva with relatively small Claparède's organ, retracted under simple tectum; protective scale-like seta 1c not discerned and no setiform homologue present. Setal ontogeny (I-III in larva, I-IV in nymphs): larva (2-1-2) or (2-1-1, with 3a absent); protonymph (3-1-2-1); deutonymph and tritonymph (3-1-2-2), with second pair on epimere IV (4a) appearing medially, near 3a; setae 1a, 2a, 3a, 4a on plicate medial cuticle, others on epimeral sclerites (see Remark 15). Coxisternal setae smooth, flexible, finely attenuate, collared (Fig. 8E).
Anogenital region — Genital valves of nymphs relatively broad, all setae aligned near medial margin; setation 1-3-5 (proto- to tritonymph) but may be variable (see below). Genital papillae flattened, as in adult. Aggenital seta (first formed in deutonymph) inserted just medial to patch p.ag. Paraproctal valves relatively long; atrichous in first three instars, without setal vestiges; variation in anal setation of tritonymph as per adult.
Gnathosoma — Mostly as in adult. Subcapitulum with laterally incomplete labiogenal articulation in all juveniles. Palp setation complete in larva, without noticeable change in nymphs or adult.
Legs — Leg shape changing during development: segments more tubular, simply constructed in larva and protonymph; femora and tibiae (especially III and IV) becoming more elongated and legs overall approaching adult shape with successive instars, but highly modified shape of trochanters III and IV attained only in adult. All tarsi with well-formed ventral guide tube for tendon of claw-depressor muscle, proportionally slightly longer than in adult, reaching proximally about 3/4 length of tarsus; ambulacral claw as in adult. Porose organs not internalized as tracheae or saccules; present as large porose areas on adaxial face of all femora (Fig. 12I), absent from trochanters III and IV. Seta d of genu IV of normal form and size; d on other genua and all tibiae small or minute, usually inconspicuous, coupled with respective solenidion. Most setae of basal four segments collared (Fig. 12H), unlike adult setae: exceptions (no collar) include enlarged setae, such as d, l′ on femora I/II of P. spooneri n. sp. and v″ on tibiae I/II of both species. Setae of tarsi generally similar in form to those of adult, except pair (ft) collared. Leg IV setation in protonymph 0-0-0-0-6: seta pv″ absent from normal tarsal complement.
ZOOBANK: 32E180E2-8CB2-4BED-8FF9-C963057CB266 ![]()
Diagnosis — With traits of Paraquanothrus n. gen. (above). Adult. Total length 766-912 μm. Cuticle moderately wrinkled dorsally and laterally; sclerites weak, easily deformed, circumgastric scissure often not well discernible from weak notogaster and ventral plate. Notogaster with distinct central dome and surrounding rim, without posterolateral tectum; setation consistently unideficient (15 pairs). With variably developed vertical ridge posterior to leg acetabulum I and distinct epimeral tooth posterior to II; acetabulum III distinctly closer to II than to IV. Leg IV (claw not included) 0.63-0.71 times body length. Trochanters III, IV and all femora with stigma and trachea. Leg tarsi without iteral setae; seta d of femora I, II inserted in proximal half of segment, d and l′ similar to other setae in form and direction. Juveniles. Hysterosoma with integumental plications finely spaced; leathery patches indistinct, pale in preserved specimens. Larva with gastronotic seta h3 and coxisternal seta 3a present.
Etymology — The species epithet honors Dr. Tim Graham, an expert on the biology of rock pool and pothole microhabitats of the Colorado Plateau, who first discovered the species and studied its biology, and who provided most of the material studied herein.
Type locality — USA; Utah; Grand County, ca. 1 km northeast of Moab; lat/long 38.583, -109.535; ca. 1460 m a.s.l.
Material examined — The following were collected from the type locality (1991, 2007, 2015) by Dr. Tim B. Graham from sediment in small, shallow rock-pools (‘pans’, 1 m or less in diameter) on barren ‘fins’ of Navajo sandstone. The type-series (51 adults, preserved in alcohol) includes: the holotype female and five paratypes, collected 5-VII-2015 and deposited in the USNM; 10 paratypes collected 16-III-2007 and deposited in the CNC; five paratypes collected 16-III-2007 and deposited in the INPA, and 30 paratypes (four from 2015, 26 from 2007) retained by the first author. Non-type material from the type locality includes 20 nymphs (from 2007, in alcohol) divided between the USNM and CNC. The first author retains the following from collections on 17-XI-1991 and 16-III-2007: 27 alcoholic and 23 dissected, slide-mounted adults; 40 alcoholic and 30 slide-mounted juveniles.
Other studied material (all collected by Tim Graham from small aquatic pans on rock surfaces and retained by first author) is also from Utah: Garfield County, Glen Canyon National Recreation Area, Highway 95, west of mouth of Dirty Devil River, 26-II-1993 (12 adults, three juveniles); Sevier/Emery County border region, route I-70, 1 km W of mile marker 94, 5-III-1993 (six adults, three juveniles).
Adult (Figs 1-6, 16A-C)








Dimensions, proportions — Total length 766-912 μm (mean 824, n = 36); maximum width 466-553 μm (mean 500, n = 36). Females slightly larger than males but with much overlap: female length 776-912 μm (mean 837, n = 10); male length 766-824 μm (mean 792; n = 10); female maximum width 485-553 μm (mean 516, n = 10), male width 466-504 μm (mean 481, n = 10). Maximum notogastral width in dorsal view about 1.6-1.7 times that of prodorsum.
Integument, setae — Cuticle at most weakly sclerotized, easily deformed. Fully mature individuals black in life when dry, shiny and iridescent under water (Fig. 16A, B; Supplemental video) due to reflective plastron; fading to light brown after preservation (Fig. 16C). Teneral individuals (prior to darkening of cerotegument): yellow-orange in life due to show-through of body contents; fading to pale yellow or cream-colored after preservation. Prodorsum, notogaster and margin of ventral plate with irregular wrinkles and low undulations of various strength and direction, as indicated below; variable among individuals. Cup of apobasic body setae longer than wide (ca. 7 x 5 μm for notogastral setae; Fig. 4B).
Prodorsum (Figs 1A, C, 2A, C, 16C) — Cuticular creases and undulation irregular and somewhat variable in position and strength; orientation of individual creases generally transverse in anterior half, vaguely longitudinal in lateral region but with well delineated longitudinal pair often forming H-pattern with most posterior transverse crease; usually with pair of angular creases running posterolaterally to bothridial region. Central cheliceral retractor muscles inserting without conspicuous muscle sigilla. Bothridial seta (Fig. 1D) directed dorsolaterally; 85-90 μm, setiform, straight or very weakly curved; isodiametric in middle third, narrowing slightly in basal third, distal region tapered either gradually or abruptly to acute tip; pale basally, becoming light brown in distal half but without apparent cerotegument; with barbs inconspicuous, minute in distal third to half, sparser or absent in basal half. Seta in present or absent (see Notes, below, for details): if present, formed as alveolar vestige or as normal, thin seta, ca. 20-30 μm, inserted ca. 30 μm medial and slightly anterior to bothridium. Setae le and ro smooth, finely attenuate: le ca. 30-40 μm, mutual distance slightly greater than length (Fig. 1A); ro similar, but mutual distance and distance from rostral margin both ca. 20 μm. Seta ex usually absent (see Notes); if present, formed as alveolar vestige (Fig. 4A) or as normal, fine seta, ca. 20-25 μm, inserted ca. 25 μm ventral to bothridium.
Notogaster (Figs 1A, C, 2A, C, 16C) — Outline nearly elliptical in dorsal view. With low, convex central dome, somewhat flattened posteriorly in contracted specimens; dome surrounded by flatter rim, usually well delineated but not separated from dome by scissure, less distinct in gravid females. Band of dorsoventral muscles inserting at junction of dome and rim in lateral region, but without conspicuous sigilla (i.e., typical circumgastric band not evident). Dome margin with vaguely circumferential creases, forming irregular reticulation; anterior half of dome featureless or with inconspicuous undulations or subsurface patterns, sometimes with one to several longitudinal creases or weak ridges (Fig. 16C); posterior half of dome with well-defined and generally transverse creases. Rim with creases strongest posteriorly, oblique or vaguely radial in orientation. Lenticulus ca. 40-50 μm long, elliptical to subrectangular, width about 0.6 length. Submarginal band of desclerotization (ascleritic band) not evident, or only weakly discernable between seta c3 and lyrifissure im. Notogastral margin without tectum, but folded margin may appear darker in transmitted light. Setation unideficient: 15 pairs of setae consistently present, all but d-series inserted on rim, with l- and h-series aligned longitudinally. Mutual distance of pair dm clearly larger than that of da and about twice that of dp. All setae thin, attenuate, straight or slightly curved, without noticeable barbs; length 22-39 μm; c3, la, lm, da, dm longest, h1 shortest, others intermediate. Lyrifissures and opisthonotal gland opening (gla) also on rim: usually im, gla, ih and ips in nearly straight, uninterrupted line, almost equally spaced; seta lp usually inserted posterior to ips, less often between ih and ips. Lyrifissures relatively inconspicuous, small, 8-13 μm long, ih shortest, im usually oriented vertically or obliquely. Circumgastric scissure hardly distinct from weakly sclerotized notogaster and ventral plate; distinguished mostly by slight, variable folding of notogastral margin over prodorsum and ventral plate.
Podolateral and coxisternal regions — With variably developed crease between acetabula I and II, appearing like vertical ridge (vr) when well-developed; epimere II projecting laterally as distinct small tooth (et) posteroventral to acetabulum II (Figs 1B, C, 4C), with inconspicuous carina extending dorsally from tooth (Fig. 3C). Sejugal stigma opening just posterior to tooth. Acetabulum III anteriorly displaced: distance from IV about four times as far as from II (Fig. 1C). Epimeres with neither clearly marked muscle sigilla nor distinct borders that extend beyond apodemes. Epimeral setae similar to those of notogaster in form and mostly in size, except setae of epimere I slightly longer (up to 55 μm); mutual distance of pair 1a little more than half that of pair 2a.
Anogenital region — Area between genital and anal plates smooth; lateral region with oblique folds or undulations. Genital plate with several weak oblique undulations; genital setae aligned on thickened medial margin, simple, attenuate; aggenital seta similar. Vertical wall of preanal organ flat, inconspicuous. Anal plate smooth except for vague, broad oblique undulations anteriorly and posteriorly; usually with two pairs of setae, an2 usually slightly longer than an1; observed variation 0-3 setae per valve, often asymmetrical (see Notes, below). Adanal setae nearly equally spaced, level of ad2 insertion clearly anterior to posterior margin of anal plate. All setae of anogenital region apobasic, ca. 25-35 μm. Lyrifissure iad small (ca. 10 μm long), inconspicuous. Paired lobes of ovipositor usually with one additional distal seta (five on each lobe), rarely also with additional proximal seta (six total), and rarely without additional setae (four total). Spermatopositor with five pairs of setae (ψ2 absent).
Gnathosoma — Labiogenal scissure of subcapitulum incomplete, stopping variably between levels of genal setae a and m (Fig. 5C). Hypostomal seta (h) smooth or with several minute barbs, finely attenuate, flexible (ca. 50-60 μm); genal setae similar in form, a (ca. 35-40 μm) slightly shorter than m (ca. 45-50 μm). Postpalpal seta ca. 8 μm. Palp (Figs 5D, 6C) with setae on basal segments finely attenuate; femoral setae thin, nearly smooth, inf (ca. 25 μm) slightly longer than sup; setae of genu and tibia sparsely barbed. Tarsal seta cm (ca. 10 μm) shorter than height of segment, baculiform, often with single barb on outer curvature; acm (ca. 7 μm) acute. Seta lt″ directed dorsodistally, spiniform or baculiform, lt′ erect, inserted more distally, near midline, usually thinner and more tapered; both ca. 5 μm. Eupathidial pair (ul) spiniform, ca. 3-4 μm. Seta vt″ isodiametric, blunt, ca. 6 μm; vt′ slightly tapered, ca. 9 μm; seta su acute to spiniform, ca. 7 μm; all three smooth, straight. Solenidion ω short (ca. 5 μm), baculiform. Chelicera (Fig. 5E) ca. 170-180 μm long; seta cha (ca. 40 μm) often bent near base to lie nearly parallel to surface, relatively thick, mostly isodiametric or weakly spindle-shaped, tapering in distal fifth to acute tip; chb (ca. 20 μm), acuminate. Terminal bulb of trachea within chelicera about 3-4 times tracheal diameter (Fig. 6N).

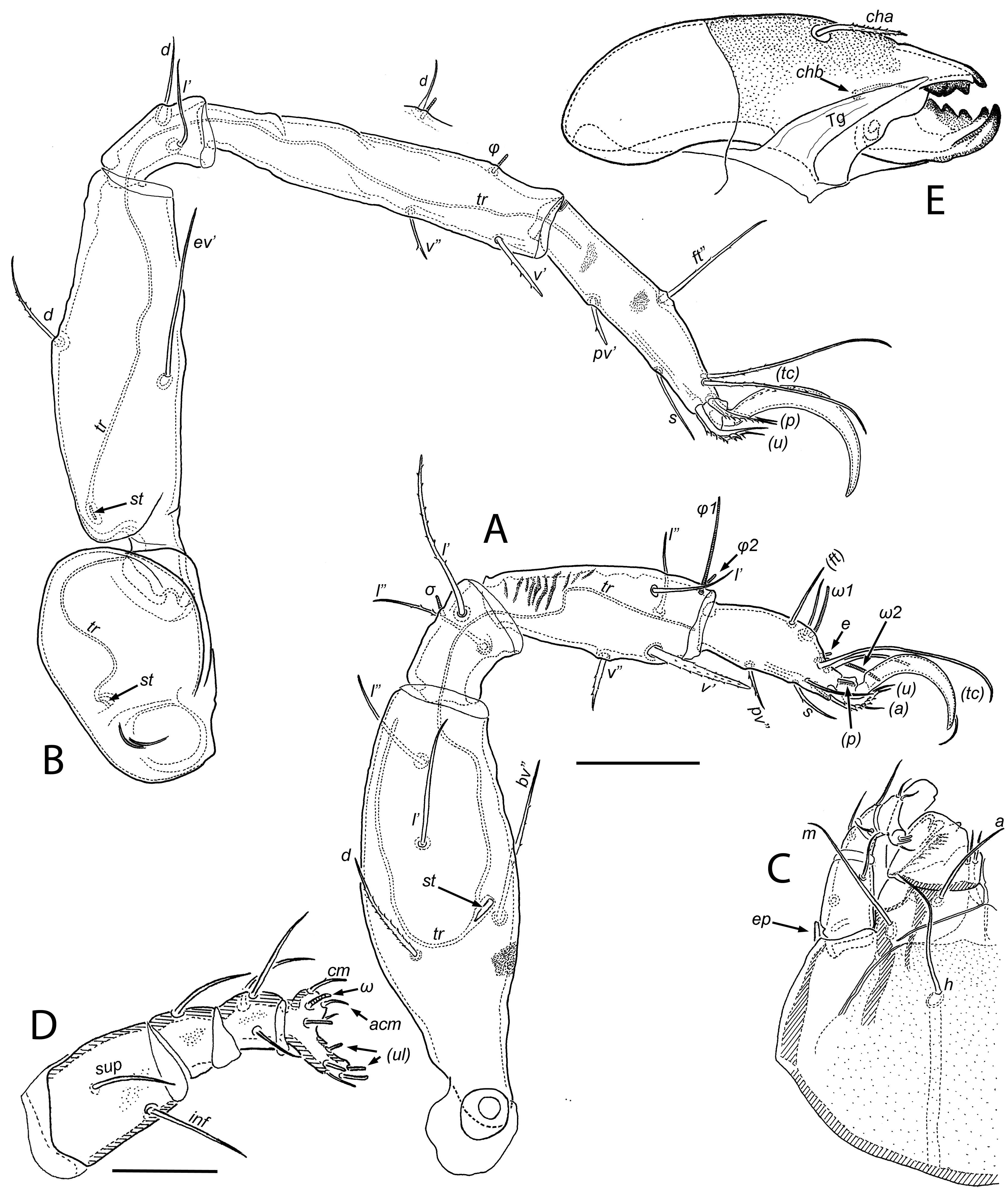



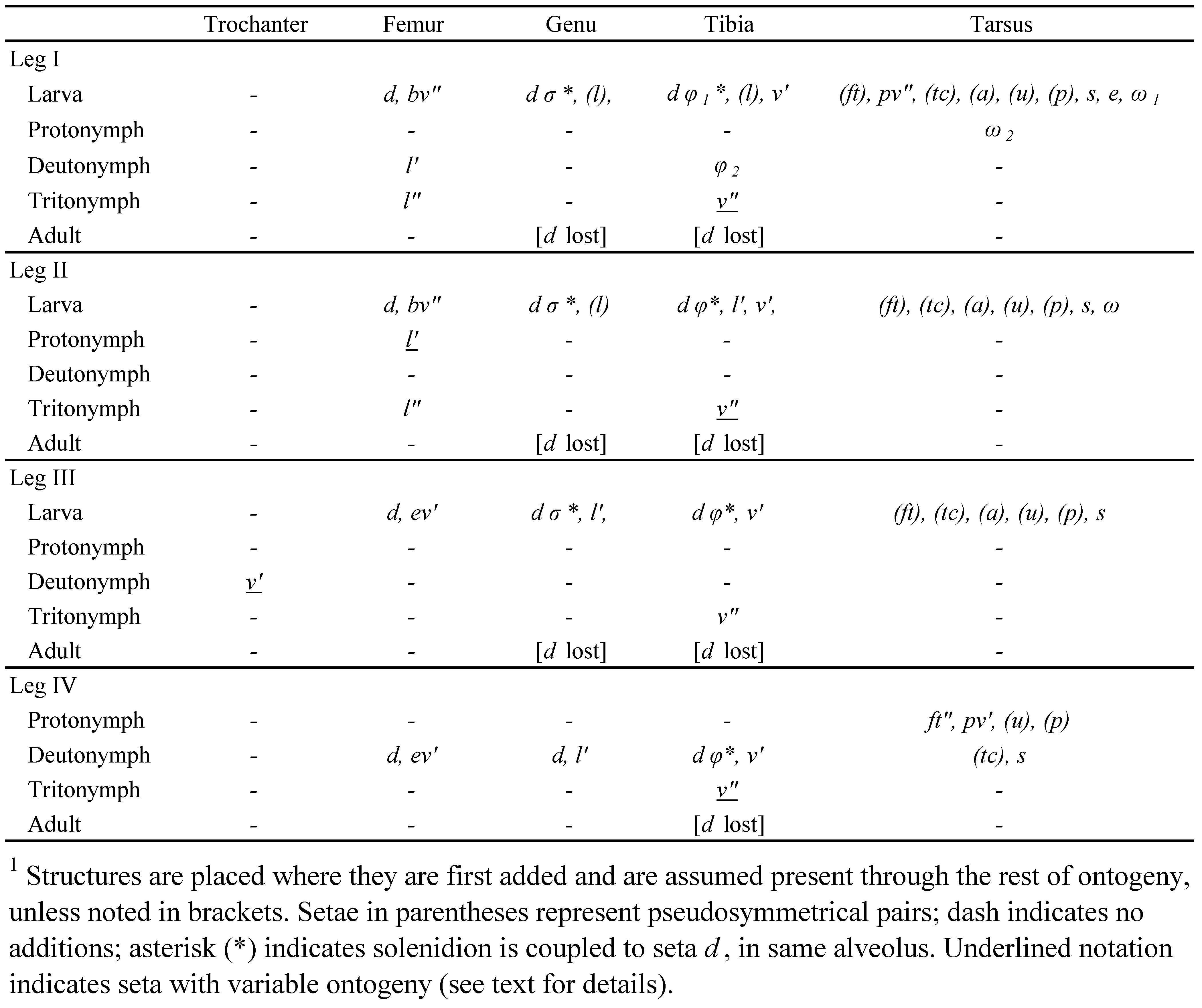
Legs (Figs 5A, B, 6A) — Leg IV (claw not included) 0.63-0.71 times body length; other leg lengths relative to IV in one measured female: I (0.74), II (0.68), III (0.82). Leg IV with tibia and tarsus about five times longer than high in lateral aspect; tarsi I and II highest at mid-length, slightly less than maximum height of respective tibia. Tibiae with cuticle slightly undulating, sometimes with vague vertical ridges proximally; other segments mostly smooth. Porose organs developed as tracheae, terminating without distinct bulb (Fig. 6D, F), ending bluntly or gradually swelling to double diameter. Femora I, II with stigma (st) in small pit low on adaxial face, opposite seta bv″, leading to flat, dark (cerotegument-lined) vestibule into which two tracheae open (Fig. 5A, 6G): posterior trachea short, remaining within segment, looping dorsally, ending variably at about 2/3 or 3/4 length of segment; anterior trachea much longer, running ventrally through femur, centrally through genu, ending variably between proximal quarter of tibia to mid-tarsus (Fig. 6D). Femora III, IV with slit-like stigma proximally, high on adaxial face, just distal to basal stalk (Figs 5B, 6H), opening into short, dark, simple vestibule with single trachea; latter running centrally through femur, genu and tibia, ending variably between mid-tibia to mid-tarsus; rarely with very short second trachea branching off near stigma. Stigma of trochanters III and IV similar to that of respective femur; trachea looping entirely within segment (rarely with additional small basal lobe extending from vestibule).
Setal counts (I-IV; homologies in Table 1 and Fig. 5A, B): trochanters 0-0-1-0, femora 4-4-2-2, genua 2-2-1-2, tibiae 4-3-2-2, tarsi (including small famulus) usually 13 -11-11-9. Seta l″ present on femora I, II; seta l′ absent from tibia III; iteral setae absent from all tarsi. Setae of femora and genua distinctly apobasic, those of tibiae and tarsi only weakly so. Tarsus I with proral setae (p) spiniform, ca. 6 μm; seta s blunt, with one or two barbs. Famulus peg-like; small (ca. 3-4 μm), inserted far distally, at base of tarsal stalk, at same level as or slightly proximal to level of tc′; internal root long, relatively conspicuous (Fig. 6D, E). Tectal setae (tc) of tarsus IV not surpassing claw, with weak or no distal thickening or curl; usually seta ft″ of tarsus IV and often pair (ft) of tarsus III (but no other leg setae) with small, inconspicuous dark basal collar. Antelateral setae (a) finely attenuate, not curled on any tarsus. Seta v′ of tibiae I, II thick, sub-spiniform, distinctly barbed. Seta d on femora I, II attenuate, barbed, distally directed, inserted in proximal third of segment, slightly proximal to level of tracheal stigma; l′ with similar size and form but more weakly barbed. Seta d of femora III, IV similar in form but variably inserted: ranging from near middle of segment, at about same level as ev′, to distal third, well separated from ev′. Distribution and form of other setae on legs I and IV shown in Fig. 5A, B; those of II, III mostly similar to I, IV, respectively. Solenidion σ of genua very small: I, II (8-10 μm), III (6 μm). Solenidion φ of tibiae: II, III (14-17 μm), IV (6-9 μm). Tibia I solenidion φ1 elongated (ca. 30-35μm), slightly tapered (ceratiform); φ2 (6-9 μm) inserted directly abaxial to φ1. Tarsal solenidia very slightly tapered. Tarsus I with ω1 (20-25 μm) just distal to seta ft″ in shared alveolus; solenidion ω2 (25-27, rarely 20 μm) with variable position on abaxial face, midway between ft″ and tc″ or closer to one of them, sometimes very near famulus. Tarsus II ω shorter (15-16 μm).
Juveniles (Figs. 7, 8, 17A-B)




Dimensions — Mean (range; number of individuals measured) total length of: larva 324 μm (281-367; n = 7); protonymph 453 μm (436-495; n = 10); deutonymph 615 μm (582-650; n = 10); tritonymph 756 μm (679-805; n = 10).
Integument — Strong pigmentation of cerotegument not retained after preservation: major sclerites (prodorsum, subcapitulum, legs) only slightly darker than soft body cuticle; leathery patches of hysterosoma only weakly pigmented, most distinguishable by weakened plication (Fig. 17B).
Prodorsum — Basal region of aspis, other than hyaline region, colored similarly to rest. Setae (except bs) with distinct basal collar. Mutual distance of pair le more than twice that of pair ro. Bothridial seta (bs) of larva thin, distally tapered, smooth or nearly so (Fig. 8D); some protonymphs similar, but bs of nymphs usually like that of adult, or slightly incrassate distally.
Gastronotic region — Somewhat hump-backed in lateral view, (Fig. 17B), with uneven curvature. Posterior margin of larva rounded to truncate in dorsoventral view (Fig. 7A), but nymphs slightly (Fig. 17A) to strongly bilobed posteriorly (Fig. 7E, F). Plication relatively fine, tritonymph with ca. 13-15 transverse plicae between setae c1, d1; with complex pattern in region bounded by setae dm, dp, lm, lp (Fig. 7E). Setae thin, flexible, finely attenuate, smooth, most with long basal collar (Fig. 8F). Setation unideficient throughout development: larva with typical 12 pairs, h3 closely adjacent to paraproct (segment PS), at about mid-length (Fig. 7B); nymphs with typical 15 pairs, p1-p3 added in protonymph in normal positions.
Venter (Fig. 7B-D, F) — Coxisternal and anogenital setae distinctly collared; seta 3a present in all instars. Genital valves of nymphs collectively elliptical to obovate. Coxisternal and anogenital setae distinctly collared. Anal setation of tritonymph highly variable: each valve with 0-3 setae, usually asymmetrical (see Notes for details).
Gnathosoma — Labiogenal articulation very short, reaching laterally only to level of genal seta a. Genal and hypostomal setae of subcapitulum usually with well-formed collar.
Legs — Setal and solenidial ontogeny given in Table 1. Seta d of femora I and II of normal form, attenuate, inclined toward surface, inserted about mid-length (unlike adult). Seta d of genua and tibiae relatively small but distinctly setiform: thin, flexible, collared. Proral setae (p) of tarsus I minute, eupathidial in all juveniles; tectal setae with form and relative size as in adult. Genual solenidion σ of larva baculiform, minute (ca. 2-3 μm), shorter than collar of closely coupled seta d; d ca. 10-12 μm; coupling similar in nymphs, except σ proportionally slightly longer, about one-third length of seta (equal to setal collar). Solenidion φ of tibiae II, III baculiform in larva, less than half length of closely coupled seta d (equal or slightly longer than collar length of d; Fig. 8G); coupling similar in nymphs, except φ proportionally slightly longer, about half length of d (2-3 times length of d collar). In nymphs, d-φ coupling of tibia IV similar to that of d-σ on genu of other legs. Tibia I of larva with φ more elongated, but isodiametric, about equal in length (ca. 15 μm) to closely coupled seta d. In all juveniles, solenidion ω1 of tarsus I coupled with seta ft″ in same alveolus; ω1 baculiform in larva, about half length of ft″; very slightly tapered (ceratiform) in nymphs, about equal to ft″ in length. Solenidion ω2 of all nymphs similar to that of adult.
Egg (Fig. 6J-L) — 280-295 μm long, elliptical, ca. 1.5-1.6 times longer than wide (n = 9); colored orange in life.
Variation in body setae – Certain aspects of setation are unusually variable. The interlamellar seta has an overall frequency in adults (n = 80) of 39% (including presence only as alveolar vestige in 3/31 occurrences). Of 40 individuals examined, it was present bilaterally (including vestiges) in 11, present unilaterally in nine (three on right, six on left), and absent bilaterally in 20. The exobothridial seta is less common, with an overall frequency (n = 74) of 15% (as vestige in 2/11 occurrences). Of 37 adults examined, it was present bilaterally in none, present unilaterally in 11 (two on right, nine on left), and absent bilaterally in 26. There was no obvious correlation between the presence or form of interlamellar and exobothridial setae, and juveniles invariably had both setae. Of 30 adults examined for anal setation, most were symmetrical, with two pairs on each side. Variations were mostly asymmetrical: one individual had a single seta on each plate, but it had the position of an1 on one side and an2 on the other; three individuals lacked one seta unilaterally and four had a third seta unilaterally. Tritonymphs seem more variable than adults: of 14 anal valves (seven individuals) examined only half had two setae, four had a single seta, two were glabrous and one had three setae; only two tritonymphs were symmetrical (1/1, 2/2). Genital setation varied only among juveniles: one protonymph (of seven examined) had two setae unilaterally; one deutonymph (of six examined) had four setae unilaterally.
Variation in leg setae – Most accessory setae (i.e. those post-larval on I-III, post-deutonymphal on IV) first appear as indicated in Table 1, but the following vertitional variation was observed. Trochanter III: v′ present on three of 14 protonymphal legs III examined. Femur II: l′ absent from two of 14 protonymphal legs II examined. Tibia I: v″ present on six of 14 deutonymphal legs I examined (bilaterally present in one individual). Tibia II: v″ present in four of 14 deutonymphal legs I examined (bilaterally present in two individuals). Tibia IV: seta v″ absent from three of 18 tritonymphal legs examined. Less variation occurred among fundamental setae, affecting only leg IV, as follows: tibial seta d retained on three of 16 adult legs IV examined (bilaterally present on one specimen; Fig. 5B insert); tarsal antelateral pair (a) present on one of 18 tritonymphal legs IV examined, a′ present on one of 14 adult legs IV examined; s absent from one different leg. Two developmental anomalies were observed: v′ was doubled on one adult tibia IV (the same leg lacking s on tarsus) and tarsus IV was deformed on leg IV of one tritonymph, with only two setae (not identifiable) in the dorsal region.
Most of our distributional and biological knowledge of this species was published earlier by Norton et al. (1996b) and Graham and Norton (1998), and is summarized here. The general ecology of the ephemeral environment was reviewed by Chan et al. (2005). In these papers the mite was considered either an undescribed species of Aquanothrus or a member of a new ameronothrid genus near Aquanothrus.
The known geographical distribution of P. grahami lies entirely within the arid to semiarid Colorado Plateau, at elevations from 1,060-1,980 m a.s.l., with most records from southeastern Utah (Graham and Norton 1998, their Fig. 2). There are no records from New Mexico, and only several from Colorado (Mesa and San Miguel counties) and Arizona (Navajo county), but these regions were less intensely sampled. The substrate is exposed, barren, fine- to coarse-grained aeolian sandstone—various cross-bedded formations of terrestrial origin—of Permian to Cretaceous age. Mites occur in shallow weathering depressions, or pans (Fig. 14A-C), that temporarily hold rainwater, typically for several days but for as little as several hours in summer. Measured inhabited pans have been from 0.5 to 10 cm depth, and most have had a surface area of 5,000 cm2 or less; pans as small as 25 cm2 have supported mites. During drying, as water level touches them, they burrow into the sand substrate, but there is no obvious correlation of mite presence with sediment depth.
Under dry conditions, mites are immobilized in the lightly cemented calcareous substrate. If artificially freed (see Supplementary video), they may remain immobile or become active; we suspect that the latter occurs if drying was relatively recent. When undisturbed dry substrate is wetted, activity is resumed almost instantly. The dense cerotegument and non-porous cuticle, combined with invaginated respiratory surfaces, appear to be highly effective at reducing water loss for weeks or months. Ad hoc laboratory observations suggest that mortality increases over time when immobilized in dry sand, but the relative importance of desiccation and starvation is unknown. Some individuals have survived one year in this state, which suggests a metabolic slow-down. Most mortality probably is abiotically induced; we saw no evidence of internal parasites, and the faunally-depauperate pans seem devoid of other small predators. The latter probably has permitted the evolution of the soft procuticle in P. grahami, with the concomitant conservation of nutritional resources that would be needed for defensive sclerotization. As described below, the congener P. spooneri n. sp., which appears to live in a biologically more complex microhabitat, has a more sclerotized procuticle and even a notogastral tectum, features that we consider defensive traits.
Tightness of the cerotegument and non-porous procuticle in P. grahami also may be responsible for the unusual length of time it takes for mites to succumb to immersion in 95% ethanol: in several tests using 5-6 individuals each, most were still active after 10 minutes of immersion, many were still mobile after 15 minutes, and it took almost a half-hour for all to be unresponsive.
Under water in small dishes, mites seem clumsy and unstable walking on the substrate (see Supplemental video), and we suspect this is due at least in part to the buoyancy added by the plastron. We do not know how much time mites spend on the surface of the substrate in nature, but in windy conditions they often can be seen helplessly caught up in water eddies at the edge of the pan; this might result in passive downslope dispersal if the pan overflows (cf. Vanschoenwinkel et al. 2008b).
Both adults and juveniles appear to be opportunistic feeders. Fecal pellets and gut contents of preserved specimens are heterogeneous (Fig. 15A-F): included in a mostly amorphous matrix are fungal spores, hyphae, diatoms, fragments of brown organic matter of unknown origin and even fragments of what appears to be plicate cuticle of juveniles. There is also much inorganic material (small birefringent or opalescent mineral particles), which could be ingested if mites were feeding heavily on surface-growing organisms with their unspecialized mouthparts.
Mites also eat orange bdelloid rotifers (Philodina sp.; see Supplementary video) that inhabit the pans; mite body contents and eggs (Fig. 6K) appear to retain the orange pigment, a unique color for internal structures of oribatid mites. The rotifer feeding organ (ramate mastax) is easily identified among gut material (Fig. 15F), and in a few instances these have comprised an entire food or fecal bolus. Feeding has not been observed directly, but mites seem too awkward on the surface to capture active rotifers. Perhaps they encounter and engulf rotifers as they burrow through loose sand, or engulf them soon after wetting, during the several minutes before the desiccated rotifers rehydrate and become active. However they are captured, rotifer density may be affected by the mites: in natural pans without mites, the rotifers can form dense orange patches, easily visible to the naked eye, which are not seen in mite-inhabited pans.
Females develop a maximum of six eggs, vertically-oriented, three in each oviduct (Fig. 6J). Even teneral females may contain eggs, the deep orange color of which is conspicuous in the pale yellow-orange body.
ZOOBANK: 2A262155-A209-4824-9724-4C539A518082 ![]()
Diagnosis — With traits of Paraquanothrus n. gen. (above). Adult. Total length 645-737 μm. Cuticle modestly sclerotized, mostly smooth, without folds, wrinkles or ridges, except prodorsum with distinct pair of carinate costulae. Notogaster with vague central dome and rim; border with distinct tectum in posterior half, covering well-defined, circumgastric scissure; setation 13 pairs (lacking two setae in p-row). Without vertical ridge or epimeral tooth posterior to acetabula I, II; leg acetabulum III about equidistant between II and IV. Leg IV (claw not included) 0.50-0.55 times body length. Trochanters III, IV with concave porose area, all femora with conspicuous saccule. Iteral setal pair present on tarsi I, II, it″ on tarsus III (it′ absent), no iteral setae on IV; seta d of femora I, II inserted in distal half of segment, d and l′ conspicuously straight, rigid, erect. Juveniles. Hysterosoma with integumental plications relatively widely spaced; leathery patches distinct, dark even in preserved specimens. Larva lacking gastronotic seta h3 and coxisternal seta 3a, both added in protonymph.
Etymology — The species epithet honors Dr. John D. Spooner, Distinguished Professor Emeritus at the University of South Carolina, Aiken. An expert in entomology and on the biology of Heggie's Rock, he discovered this species, studied aspects of its biology and provided specimens for our study.
Type locality — U.S.A., Georgia, Columbia County, near Appling; Heggie's Rock Preserve, lat 33.5472, lon -82.2569, elevation ca. 100 m a.s.l.
Material examined — The holotype female and 48 paratype adults, along with 73 non-type juveniles, were collected from the type locality on 27 January 1997 by John D. Spooner, from coarse sand lining shallow weathering depressions in a granite outcrop that support ephemeral rain pools. The holotype, five paratype adults and five juveniles are deposited in alcohol in the mite collection of the USNM. Five paratype adults and five juveniles in alcohol are deposited in the CNC and the Georgia Museum of Natural History (Natural History Building, University of Georgia, Athens, Georgia). Two paratype adults are in INPA, and the remaining paratype adults (eight in alcohol, 28 dissected and slide-mounted) and 63 juveniles (alcohol and slide-mounted) are retained by the first author.
Adult (Figs 9-11, 12A-E, 16D)





Dimensions, proportions — Total length 645-737 μm (mean 678, n = 22); maximum width 349-407 μm (mean 372, n = 23). Genders similar in size (see Notes): female length 645-737 μm (mean 681, n = 12); male length 655-698 μm (mean 678; n = 9); female width 349-407 μm (mean 373, n = 12), male width 345-378 μm (mean 372, n = 9). Maximum notogastral width in dorsal view about 1.4 times that of prodorsum.
Integument, setae — Fully mature preserved individuals chestnut to dark brown (Fig. 16D); teneral individuals orange-brown, often with darker legs and genital plates. Cuticle modestly sclerotized, shiny, without conspicuous folds or wrinkle-like ridges, except as noted. Surface of cerotegument variously foveo-reticulate or nodulate to verrucose, according to body region (see below). Cup of apobasic setae ca. 4 μm wide, 6 μm long for notogastral setae (Fig. 10E).
Prodorsum (Figs 9A, C, 10A, 16D) — With distinct pair of longitudinal, carinate costulae in mid-region, mutual distance about equal to length; intervening cerotegument weakly foveo-reticulate. Costulae connected near base by less distinct transverse crease, from center of which pair of similar creases directed posterolaterally, outlining triangular basal region of prodorsum. Distinct proximocentral patch of muscle sigilla (cheliceral retractor muscles) visible in transmitted light. Dorsophragma often with scalloped margin, that of pleurophragmata smooth or nearly so (Fig. 10A). Rostral margin slightly concave distally on each side, leaving slightly projecting medial lobe (Fig. 9A, insert) with rostral seta inserted at each corner; with or without weak vertical crease running dorsally from near setal insertion, giving rostrum narrowly truncated appearance (Fig. 9B). Bothridial seta (bs; Fig. 9D) ca. 100-110 long, isodiametric in mid-region, tapering proximally and distally; distal half darkened, with small, conspicuous barbs; basal half unpigmented, smooth. Interlamellar seta (in) inserted medial and slightly anterior to respective bothridium, mutual distance of pair only slightly greater than that of costulae; in invariably present, weakly attenuate, smooth; length variable, rarely short (ca. 10 μm), usually 30-35 μm (Fig. 9D). Exobothridial seta (ex) variable, formed as a simple small seta (up to 25 μm), a vestige, or absent (see Notes, below). Lamellar seta (le) thin, attenuate, smooth or with several minute barbs, ca. 25-35 μm; pair divergent, mutual distance about 1.5 times their length. Rostral seta (ro) of similar length but usually slightly (up to twice) thicker, with more conspicuous barbs; pair divergent, with mutual distance about equal to setal length, distinctly less than that of le.
Notogaster (Figs 9A, C, 16D) — Outline weakly ovate in dorsal view, slightly concave between levels of setae c3 and la, resulting in weakly defined humeral region. With poorly defined central dome and surrounding rim, separated by shallow, broad sulcus in region of circumgastric muscle sigilla (cms; Fig. 9C); best seen in cross-section (Fig. 10D) or oblique lighting (see Supplemental video). Sulcus usually not developed posteriorly, and often represented anteriorly by stronger depressions posterolateral to lenticulus (Fig. 16D, arrow). Cuticle without creases, densely nodulate to verrucose throughout; usually nodulate centrally (Fig. 10E) and more verrucose laterally; elongated verrucae short, their orientation mostly longitudinal in submarginal region, often vertical near margin (Figs 9B, C, 10B). Lenticulus elliptical to slightly obovate, ca. 40 x 30 μm. With narrow, inverted U-shaped ascleritic band (asc) sharply to vaguely defined in anterior half; transverse part running slightly anterior to lenticulus (Figs 9C, 10B): lateral arms of band running between seta c3 and lyrifissure ia, effacing posteriorly at level of lyrifissure im. Posterior half of notogastral margin with well-defined tectum, arising at level of acetabulum IV and increasing posteriorly over short distance to depth of ca. 45 μm (nt; Figs 9C, 10C); total depth of tectum maintained around posterior of notogaster, but solid limb portion narrower posteriorly (λ; Fig. 10F). Circumgastric band of muscle sigilla distinct. Lyrifissures conspicuous, 15-18 μm long, im usually longitudinal or nearly so, others vertical or oblique; ips variable in position, relative to ih and seta p3. Setation trideficient: 13 pairs of setae present, with single seta in p-row (p3). Setae lm and lp inserted slightly more medially than la, such that l-series not aligned with h-series, im and gla. Mutual distance of da, dm, dp about equal. All setae thin, acicular, with sparse, inconspicuous barbs, ca. 25-40 μm; extreme tip often broken, with seta appearing isodiametric.
Podolateral and coxisternal regions (Fig. 9B, C) — Without vertical ridge or epimeral tooth posterior to acetabula I, II: surface smooth or with at most vague vertical crease. Acetabulum III with typical position, about 1.5 times as far from IV as from II. Epimeral cuticle without nodules, but weak surface irregularities visible in oblique lighting; some muscle sigilla relatively well-marked in cleared specimens; without distinct epimeral borders extending beyond apodemes. Epimeral setae finely attenuate, smooth, ca. 40-45 μm. Mutual distance of pair 1a only slightly less than that of 2a.
Anogenital region (Fig. 9B) — Region between genital and anal plates smooth; lateral region nodulate or weakly verrucose to sub-reticulate. Genital plate mostly smooth, with or without several weak creases running obliquely posterolaterad from anteromedial corner of each plate; setae acuminate, ca. 25 μm, anterior often slightly longer (up to 38 μm). Aggenital seta ca. 30 μm. Vertical wall of preanal organ cupped, relatively conspicuous (Fig. 10G-J). Anal plate weakly nodulose-verrucose; each plate with two, sometimes three simple setae (ca 30 μm; see Notes). Adanal setae (20-25 um) unevenly spaced; ad2 posterior to margin of anal plate, ad3 frequently absent (see Notes). Lyrifissure iad distinct (ca. 15-18 μm long). Paired lobes of ovipositor each usually with typical four setae (one distal, three proximal), rarely with one additional distal seta. Spermatopositor with typical six pairs of setae (ψ2 present).
Gnathosoma — Labiogenal scissure apparently complete (Fig. 11C; observation complicated by cuticular fold). Hypostomal seta (h; ca. 45-50 μm) smooth or with several minute barbs, finely attenuate, flexible tip often broken; genal seta m similar, a slightly shorter (ca. 35 um). Postpalpal seta (ep) ca. 7 μm. Setae on basal segments of palp (Fig. 11D) with various forms: femoral seta sup acuminate, heavily barbed in contrast to thin, smooth attenuate inf; d of genu and tibia acuminate to acute, strongly barbed; pair (l) of tibia attenuate, moderately to sparsely barbed. Tarsal seta cm (ca. 15 μm) about equal to height of segment, spiniform to baculiform, often with single barb on outer curvature; acm about half as long, acute to spiniform. Setae of pair (lt) similar (ca. 5-6 μm, thin, acuminate), both erect, inserted at same level. Eupathidial pair (ul) spiniform, ca. 4-5 μm. Pair (vt) nearly isodiametric, slightly roughened, vt′ ca. 9 μm, vt″ slightly shorter; su spiniform, ca. 9 μm; all three finely roughened and slightly curved dorsally. Solenidion ω ca. 9 μm, baculiform, weakly curved. Chelicera (Fig. 11E) ca. 130-150 μm. Setae cha, chb ca. 30-35 μm and 20 μm, respectively; tracheal bulb usually expanded to about twice width of trachea.
Legs (Figs 11A, B, 12A) — Leg IV (claw not included) 0.50-0.55 times body length; relative length of other legs to IV approximately: I (0.80), II (0.75), III (0.85). Leg IV with tibia and tarsus about four times longer than high in lateral view; tarsi I and II highest in proximal third, equal to or slightly greater than maximum height of respective tibia. Cuticle of larger segments (not genua or trochanters I, II) with weak, irregular vertical or circumferential undulations and vague, sometimes finely reticulate cuticular patterns. Trochanters III and IV (Fig. 11B) with concave porose area (ca. 15-20 μm diameter) on adaxial face, dorsal to articulating stalk. Femora with conspicuously dark (from cerotegument lining) saccules of two types: I, II with elongated purse-like saccule (ca. 50-55 μm long, 7-8 wide) on adaxial face, opening via small proximal slit (Figs 11A, 12B); femora III, IV with very short (ca. 15 μm long, 7 wide) saccule opening in depression high on proximal vertical wall, just distal to basal stalk (Figs 11B, 12C).
Proximal region of leg setae colorless, strongly birefringent; pigmented outer layer (including barbs if present) not reaching setal base. Setal counts (I-IV): trochanters 0-0-1-0, femora 3-3-2-2, genua 2-2-1-2, tibiae 4-3-3-2, tarsi (including minute famulus) 15-13-12-11. Setal homologies indicated in Table 2 and Fig. 11A, B. Seta l″ absent from femora I, II; seta l′ present on tibia III; iteral pair (it) present on tarsi I-II, it″ on tarsus III; antelateral pair (a) present on tarsus IV. Most leg setae with normal insertions, not apobasic. Tarsus I with proral setae (p) eupathidial, spiniform, ca. 7-8 μm; seta s attenuate, strongly barbed on outer curvature. Famulus spiniform, minute and variable: ca. 2 μm or less, inserted closely distal to solenidion ω2 but in same or separate alveolus; internal root present but inconspicuous. Tectal setae (tc) of tarsus IV similar to those of other legs. Fastigial setae (ft) of tarsi III, IV with swollen birefringent base but without noticeable collar. Seta v′ of tibiae I, II distally attenuate, with two rows of dense barbs on outer curvature. Antelateral setae of all tarsi with distal curls (like those of tectal and iteral setae). Seta d of femora I, II erect, conspicuously thickened and barbed, tapered only in distal quarter, inserted in distal third of segment; l′ similar in form, but noticeably smaller. Seta d of femora III, IV stiff, clearly barbed, but angled distad, usually inserted in distal half or third of segment, well distal to level of thinner, attenuate ev′. Seta d of femora III, IV inserted in distal third of segment, far distal to level of ev′ insertion.
Solenidion σ of genua minute (5-7 μm). Solenidion φ of tibiae mostly similar on II-IV but of various lengths: II, (15-16 μm), III, IV (6-8 μm); φ1 of tibia I slightly tapered, longest of all solenidia (ca. 42 μm), φ2 among shortest (5-6 μm), inserted distinctly proximal to level of φ1. Tarsus I ω1 (25-26 μm) slightly tapered (ceratiform), adjacent to seta ft″ but not in same alveolus, ω2 slightly distal and abaxial to ω1, slightly thinner, more finely tapered, longer (32-33 μm), sometimes sharing alveolus with minute famulus (see above). Tarsus II ω ca. 15 μm, very slightly tapered.
Juveniles (Figs 12F-L, 13, 17C-D)





Dimensions — Mean (range; number of individuals measured) total length of: larva 306 μm (292-320; n = 2); protonymph 364 μm (340-388; n = 2); deutonymph 486 μm (456-524; n = 7); tritonymph 576 μm (543-621; n = 10).
Integument — Cerotegument as in adult, with papillae not fused, resulting in surface minutely roughened by dense, fine, dark surface granulation; that of gastronotum often with small adherent debris particles, including scattered, non-epibiotic diatoms, making small structures (e.g. cupules) difficult to observe. Dark pigmentation of cerotegument with irregular, nearly black splotches; conspicuously retained after preservation on prodorsum, subcapitulum, legs and leathery patches (Fig. 12F, G), but not on epimeral plates or anogenital valves.
Prodorsum (Fig. 13A, E) — Trapezoidal region at base of aspis dark (Fig. 17C), cerotegument with irregular, nearly black blotches underlain by porose procuticle. Setae with collar weakly developed or absent. Setal pair le relatively close together, with mutual distance little greater than that of pair ro. Seta ex unilaterally absent from some nymphs (not enumerated); both lamellar setae absent from one deutonymph. Bothridial setae similar in all instars, but barbs less conspicuous in larva.
Gastronotic region — Outline smoothly convex in lateral view, hysterosomal thickness uniform (Fig. 17D). All instars with posterior margin broadly rounded in dorsoventral view, never bilobed (Figs 13, 17C). Plication relatively coarse, tritonymph with ca. 6-8 transverse plicae between setae c1, d1; pattern simple, mostly transverse in region bounded by setae dm, dp, lm, lp (Fig. 13E). Hysterosomal leathery patches darkly pigmented even in preserved specimens, contrasting strongly with surrounding cuticle (Fig. 17D) and underlain by porose procuticle (Fig. 12F). Setae rigid, straight, acicular, or weakly curved, with small but distinct barbs. Larva with 11 pairs (h3 absent); nymphs with 13 pairs, h3 and single p-seta (p3) added in protonymph.
Venter — Coxisternal and anogenital setae often with basal collar indistinct or absent; seta 3a absent from larva, present in nymphs and adult. Genital valves of nymphs collectively diamond-shaped (Fig. 13C, D, F). Coxisternal and anogenital setae often with basal collar indistinct or absent.
Gnathosoma — Labiogenal articulation reaching laterally more than halfway between level of genal setae a and m. Genal and hypostomal setae of subcapitulum usually with basal collar weakly formed or absent.
Legs — Setal and solenidial ontogeny given in Table 2. In all juvenile instars, seta d of femora I and II as in adult: erect, thickened, barbed and inserted distal to mid-length of segment. Seta d of genua and tibiae of various forms. Seta d of gena I-III ca. 3-4 μm in larva, spiniform, in same alveolus as slightly shorter, baculiform solenidion σ, but diverging in direction (Fig. 12J); nymphs with σ proportionally similar, but seta d about twice length of σ, acicular, with distinct basal collar. In larva, seta d of tibiae II, III minute, smooth, tapered, without collar, inconspicuously fully appressed to side of small, baculiform solenidion φ (Fig. 12K), not extending beyond φ but true length not discerned; in nymphs, d not appressed to φ and about twice its length, acicular, with basal collar (Fig. 12L). In nymphs, coupling of d-φ on tibia IV similar to that of d-σ on genu of other legs. Tibia I of larva with seta d similar to that of tibiae II, III, but appressed to much longer, finely tapered (piliform) solenidion φ (ca. three times as long as d); d spiniform in nymphs, about one-tenth length of φ. In all juveniles, solenidion ω1 of tarsus I adjacent to seta ft″ but in separate alveolus; ω1 baculiform in larva, about half length of ft″; very slightly tapered (ceratiform) in nymphs, about equal to ft″ in length. Solenidion ω of tarsus II isodiametric, elongate-baculiform in larva; shorter than fastigial setae in all nymphs; isodiametric in protonymph, very slightly tapered in deuto- and tritonymph.
Egg — Elliptical, 245-318 μm long, ca. 1.6-1.8 times longer than wide (n = 7).
Variation in body setae — The exobothridial seta (ex) is highly variable in adults (n = 39 observations): it can be a thin, simple seta, 7-25 μm long (18 of 39; Fig. 9D), represented only as an indistinct alveolar vestige (15 of 39), or absent (six of 39); the form in a given individual is generally symmetrical (similar, but not identical). Of 17 adults examined: coxisternal seta 4a was absent unilaterally from one. Adults usually have two pairs of anal setae, but six of 17 adults had three setae unilaterally; tritonymphs may have a single seta and an alveolar vestige of the second (not enumerated, but symmetrical in one specimen). Adanal seta ad3 is highly variable: of 20 adults examined, it was present bilaterally in eight, unilaterally present in eight, and absent bilaterally in four (overall frequency 0.6); variation was present in deuto- and tritonymphs but not enumerated. Genital setae varied only in one deutonymph: two setae on one valve, one on the other, instead of the usual three pairs (Fig. 13D).
Variation in leg setae — In contrast to the highly variable leg setation of P. grahami, we noted only three instances that deviated from Table 2. Seta l″ was present on femur I in one of 20 adult legs examined. Seta l′ failed to form on tibia IV in two of 20 adult legs examined.
Size-variation and gender — Paraquanothrus spooneri is unusual among brachypyline oribatid mites in having males and females of nearly the same size (means 678 and 681 μm, respectively). In a few taxa, most notably many species of Oripodidae, males are distinctly smaller than females and lengths do not overlap (e.g. Travé 1959, Baranek 1981), and rarely males are slightly larger than females (e.g. the galumnid Cryptogalumna cryptodonta Grandjean, 1957; Bernini 1984). But in most oribatid mite species the size-ranges of females and males overlap somewhat, with the mean for males being clearly smaller. For example, the median male length of the podacarid species Podacarus auberti Grandjean, 1955 is about 94% that of females (Grandjean 1955), and mean length of males in several Ameronothrus species ranges from 89-94% that of females (Schubart 1975). Paraquanothrus grahami follows this typical pattern, with the male mean being about 95% that of females.








Notogastral tectum — In some oribatid mites, the soft circumgastric scissure is entirely or partially protected from predators by an overlapping tectum formed on the notogastral border. It is uncommon in macropyline groups but characterizes the ptychoid family Phthiracaridae and the synichotritiid subfamily Temburongiinae (Norton and Lions 1992). Among Brachypylina the humeral pteromorph is such a tectum, but a less conspicuous posterior notogastral tectum may be present, as well as a tectum across the anterior border; all these are most common in Poronota (Norton and Behan-Pelletier 2009), where they also usually characterize family-group taxa. The notogastral tectum of Paraquanothrus spooneri is the second to be reported among Ameronothroidea. The first was found in the podacarid species Antarcticola meyeri Wallwork, 1967; Wallwork (1967) described it as ‘a broad, free tectum with humeral lobes resembling horizontal pteromorphs anterolaterally’, but this is somewhat misleading. Based on our study of topotype specimens from Antarctica, the part delineated by dotted lines in Wallwork’s Fig. 4—from the region of seta la around the front to the other seta la—is a wide, flattened (but internally open) horizontal fold in the notogaster; unlike a pteromorph, it has rounded outer edges in cross-section, and is only marginally describable as a tectum. From that point (approximately the level of acetabulum III) posteriorly, the notogastral margin forms a broad true vertical tectum with a sharp ventral edge that runs completely around the posterior of the mite. It appears much like the convergently evolved tectum of P. spooneri (our Fig. 10C, D) except there is no solid limb at its edge. Wallwork (1967) overlooked the posterior tectum but we agree with his assessment that—despite the anterior fold and several other unusual features of A. meyeri (e.g. the loss of dorsocentral setae in the adult and lack of porose dorsal sclerites in juveniles)—the species seems well-placed in Podacaridae.
Paraquanothrus spooneri is unique among the four known ameronothroid rock pool mites in being found at a low elevation, mesic location. The type (and only known) locality, Heggie's Rock, is surrounded by a mixed forest of Quercus, Carya and Pinus, but the barren granite outcrops are microclimatically rigorous. As with the exposed sandstone depressions of the Colorado Plateau, ephemeral pools vary in their depth and longevity, and therefore their biotic communities. According to Dr. John Spooner (personal communication, 1997), P. spooneri inhabits only the shallower solution depressions, less than 1 cm depth with a coarse, sandy lining, that support growths of a rare stonecrop: elf orpine — Diamorpha smallii Britton ex Small (Crassulaceae). The mite can also inhabit unvegetated depressions where elf orpine grew in the past, and where plant fragments remain. In small dishes in the laboratory, he observed immersed mites feeding on such dead stonecrop fragments. Like P. grahami, they move slowly when in water and move into the substrate when it becomes dry. He found they can survive four weeks or more in dry sediment.Observed females carried a maximum of four eggs, two in each oviduct.
Aquanothrus montanus was described from samples of three populations in the Free State, South Africa. Two were from sediment in temporary water bodies (‘natural aquaria’) on aeolian sandstone at the top of mountains. The type locality is ‘mountain Vegkop’ (Vechtkop), which according to Dr. Lizel Hugo-Coetzee (based on collection records of the Museum) has coordinates of 28°47.36′S, 27°13.87′E (there are several mountains or hills with that name in South Africa). The other mountainous site is Thaba Patchoa (29°19.9′S, 27°05.4′E; Fig. 18H-I). The geological substrates are similar in origin and age to that of the type locality of Paraquanothrus grahami in Utah. The third sample came from a very different site: the bottom of the H. F. Verwoerd Dam (now Gariep Dam; 30°37.5′S, 25°30.4′E), 30 m below the surface. According to Dr. Louise Coetzee, who was involved with the collection, all the latter specimens came from a ‘ball of debris’ and she speculated that they might have been washed down the watershed to the dam in heavy rain, rather than representing an in-situ population.


Like Paraquanothrus species, A. montanus moves clumsily, can dig into the substrate, and survives extended dry periods. The mites feed on sediment but also on aquatic plants. There have been no further morphological studies, but mites identified as this species were part of rock pool faunas investigated in several community ecology studies. Vanschoenwinkel et al. (2007, 2008a, b, 2009) studied metacommunity structure and dynamics in sandstone rock pools on Korannaberg Mountain, also in the Free State. They found the mite to be dispersed by wind and by overflowing water, and these probably are also the dispersal mechanisms of Paraquanothrus species. Mites attributed to A. montanus (but see below) were also part of the rock pool fauna on granite outcrops in arid to semiarid southeastern Botswana, studied by Jocqué et al. (2006, 2007).
Engelbrecht (1975) viewed his South African material as representing a single species that varied greatly in body surface patterns, notogastral setation and the form of certain leg setae. However, he did not discuss these variations in the context of the three populations, nor did he identify the source for illustrated specimens or indicate correlation among the different versions of traits. In essence, he concluded that the general sympatry of the variants precluded the existence of several species or subspecies. But the total extent of reported morphological variation exceeds that of any other single known species of oribatid mite. For example, the tarsal setation in the limited number of adults from Thaba Patchoa available to us is strikingly (and consistently) different from that of the other two sites (see below), and those of the Botswana sample are different still. We think there are at least three endemic species of Aquanothrus in southern Africa, represented by the Vegkop (and Verwoerd Dam), Thaba Patchoa, and Botswana populations, and hereon we treat the latter two as representing undescribed species. Considering the highly insular nature of the habitat and the isolation-potential of mountainous environments, fine-grained speciation should not be surprising.
The morphological studies of Engelbrecht (1975) are extensive and detailed but include certain errors and omissions that hamper comparisons with other ameronothroid mites. Below, we: correct these errors based on studies of paratypes; provide new data on leg setal ontogeny; and propose a new and expanded generic diagnosis for Aquanothrus, comparable to that for Paraquanothrus, that encompasses all populations studied by us.
Engelbrecht (1975) presented illustrations (his Figs 18-21) of all four legs from one variant, which corresponds with the paratypes from the type locality at Vegkop Mountain studied by us, as well as the sample from Verwoerd Dam. He also illustrated (his Figs 22-25) the four tarsi from another variant, which consistently matches the paratypes from Thaba Patchoa studied by us. The most striking differences between variants relate to the form of tectal and fastigial setae but solenidial placement and the famulus also differ. On comparing these figures with dissected paratype specimens, we noted the problems described below; further explanations are given in reference to the complete setal ontogeny.
Engelbrecht (1975), Fig. 18 (leg I) — On the genu, the long structure drawn as a solenidion (labeled σ and provided with the traditional transverse cross-hatching) is actually seta d (it is birefringent in polarized light); solenidion σ is the short one (it is isotropic). The presence of d seems typical of the population, but not invariable (see Table 3 and below). Several problems relate to the tarsus. Seta ft″ was drawn (just distal to ?pl) but not labeled; of the two setae labeled pv, the proximal one is pv″, the distal one is the subunguinal, s. The seta he labeled ?pl is actually pv′ and is incorrectly drawn on the abaxial (= ‘antiaxial’) side (cf. the correct Fig. 19). This position of fundamental seta pv′, high on the adaxial face of tarsus I in all instars (and also tarsus II; see below), is unusual.
Fig. 19 (leg II) — On the genu, observations in polarized light showed that the long dorsal structure is seta d; solenidion σ is the short one. On the tarsus: setae labeled it′ and it″ are respectively tc′ and tc″; the fastigial pair (ft) are drawn (in the vicinity of solenidia) but not labeled; of the two setae labeled pv, the proximal one is pv″, the distal one is seta s. As for the previous figure, the seta labeled ?pl is an unusually placed pv′. It is not a primilateral seta, since these never occur on tarsus II in Brachypylina (Grandjean 1959). Also, the seta is clearly not a proximal accessory seta in the l′ row since it is present from the larva on both tarsi I and II.
Fig. 20 (leg III) — On the tarsus, the seta labeled pv″ is the subunguinal, s.
Fig. 21 (leg IV) — On the genu, the dorsal hatched structure is seta d (as in nearly all Brachypylina there is no solenidion on this segment; see below). On the tibia the dorsal hatched structure is seta d; solenidion φ is at its base, minute and not drawn. On the tarsus the single fastigial is ft″, not ft′; the setae labeled ?Ad, pv′ and pv″ are respectively pv″, pv′ and s; therefore, pair (pv) has a ′ disjunction. In the Vegkop population, no tarsal seta is added in the adult on any leg (see below).
Fig. 22 — The pair labeled it′ and it″ are respectively tc′ and tc″. The seta labeled ft′ is solenidion ω1, tightly coupled to the large ft″. The structure labeled ω2 is ft′, and the solenidion labeled ω1 is actually ω2. The small, non-eupathidial proral seta p′ is present, but hidden from view, and the unlabeled ventral seta is s.
Fig. 23 — The pair labeled it′ and it″ are respectively tc′ and tc″. The small proral seta p″ is present, but hidden from view, and the unlabeled ventral seta is s. Solenidia ω1 and ω2 are correctly labeled; they are closely coupled, but in separate alveoli.
Fig. 24 — The pair labeled it′ and it″ are respectively tc′ and tc″. The small proral seta p″ is present, but hidden from view, and the unlabeled ventral seta is s. In none of our specimens was seta a′ truncated as shown in the figure; all had a′ attenuate, similar to that of tarsus IV (Fig. 25).
Fig. 25 — The pair labeled it′ and it″ are respectively tc′ and tc″. Setae p″, a″ and u″ are present, but hidden from view, and the unlabeled ventral seta is s.

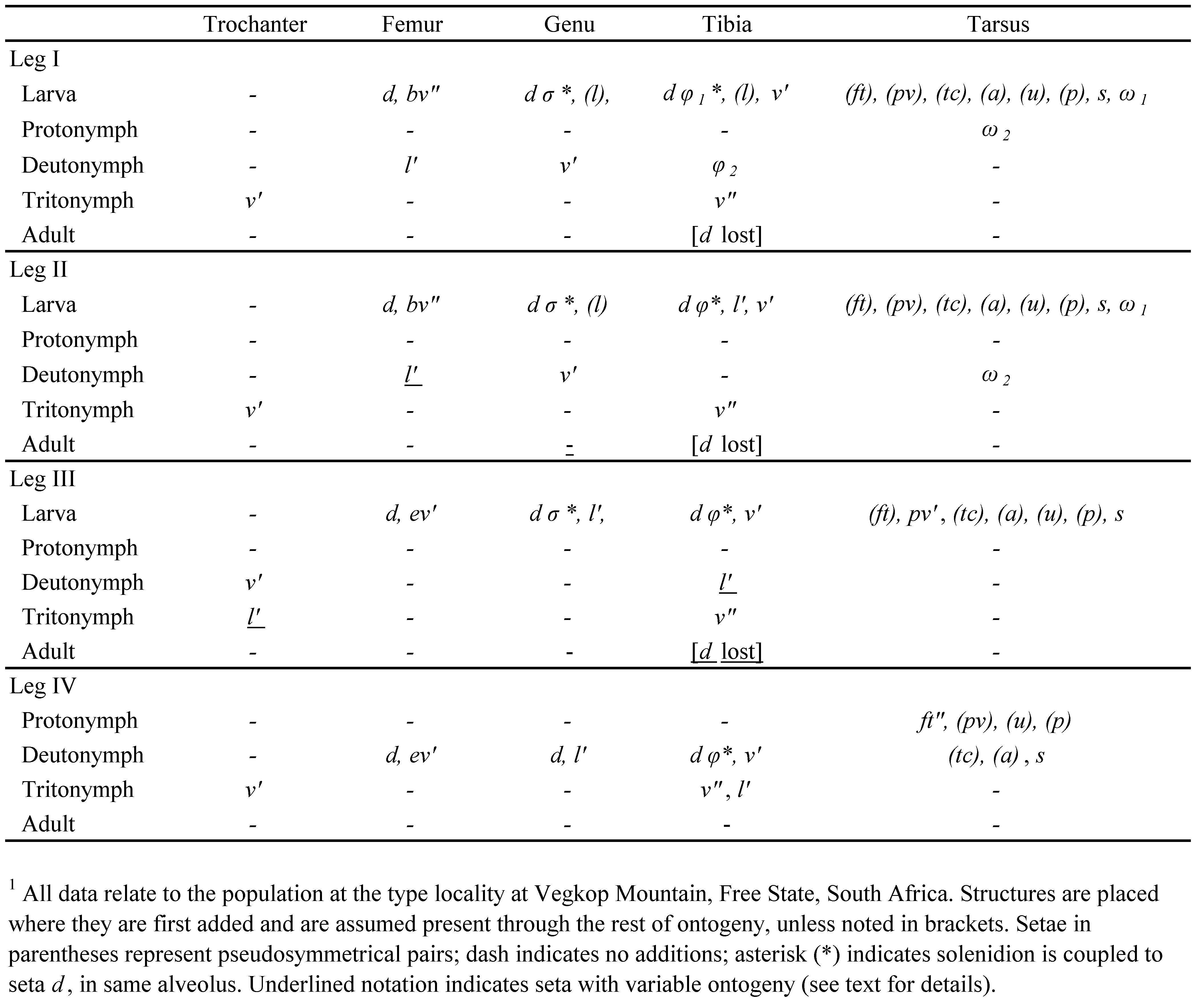
Table 3 outlines the development of leg setation for the population of Aquanothrus montanus at the type locality, Vegkop Mountain. Highlights of differences with Paraquanothrus species include: (1) the addition of seta v′ (tritonymphal) to trochanters I, II; (2) the retention by the adult of seta d on genua I-II and tibia IV; (3) the addition of v′ (deutonymphal) to genua I, II; (4) the absence of the famulus from tarsus I in all instars (but see below); (5) the normal structure of proral setae (not eupathidial) on tarsus I of all instars; (6) the close coupling of tarsus I solenidion ω1 to ω2 beginning with the protonymph (coupled with ft″ in the larva, as in Paraquanothrus); (7) the presence of fundamental seta pv′ on tarsi I-III; (8) the presence of seta pv″ on tarsus IV, giving the normal oribatid mite complement of seven protonymphal setae.
Noted variations within and differences among the three studied populations include the following. Genual seta d: of four examined paratype adults from Vegkop, one lacked seta d bilaterally from genua I and II; on these segments the Thaba Patchoa species retains seta d, but the Botswana species consistently loses d in the adult. Seta d of tibia III is highly variable in the Vegkop population, being absent from the leg illustrated by Engelbrecht (1975, his Fig. 20) and three of seven legs III of Vegkop paratypes studied by us (retained in the other four); the single dissected specimen from Thaba Patchoa lacked d bilaterally, as did the four examined Botswana specimens. In the Vegtop population seta l′ varies on three segments: it was absent from two of the four tritonymphal trochanters III examined, and also from about half those of adults; on femur II it was absent from one of four deutonymphal legs and one of four tritonymphal legs examined, but was present in all studied adults; on tibia III it was absent from two of four deutonymphal legs III examined, but present consistently in later instars.
Respiratory organs of legs — Engelbrecht (1975) neither mentioned nor illustrated the respiratory organs of legs in A. montanus: in juveniles, the trochantero-femoral system is in the typical form of porose areas, but in adults of all three species of Aquanothrus that we studied, they are invaginated as tracheae. All four femora have a conspicuous stigma and dark tracheal vestibule (similar to those of legs I and II in Paraquanothrus grahami; cf. our Fig. 6G), from which two tracheae extend: the proximal one is short and remains within the segment, but the long distal trachea reaches various distances into other leg segments, according to species. Unlike P. grahami, the stigma on each femur is high on the adaxial face, and similarly placed on all legs. Trochanters III and IV have a smaller vestibule, also high on the adaxial face, and a single short trachea.
Famulus — Mites from the type population of A. montanus at Vegtop Mountain (and those from Verwoerd Dam) lack a famulus. Such a loss is highly unusual among oribatid mites, though one of the instances is in the podacarid genus Halozetes (reviewed by Fuangarworn and Norton 2013). In the Aquanothrus species from Botswana, the famulus is represented only by an alveolar vestige between solenidia ω1 and ω2. By contrast, in the Thaba Patchoa species there is a minute, spiniform famulus that is difficult to see, inconspicuously located immediately distal to the cluster containing the massive seta ft″, the tightly coupled solenidion ω1, and the adjacent (but with separate insertion) ω2; it is not shown in Engelbrecht's (1975, his Fig. 22) illustration, which we think represents this species.
Proral setae of tarsus I — In the vast majority of oribatid mites, setal pair (p) on tarsus I are eupathidia (hollow, pale, smooth uniporous chemosensilla that are homologues of normal setae), but in all examined specimens from the Vegkop and Verwoerd Dam samples of A. montanus they are normal, pigmented, and indistinguishable from those of tarsi II-IV. Since the subunguinal seta of tarsus I is also normal, there are no eupathidia on legs of A. montanus from Vegkop, nor in the Thaba Patchoa species. These are the first such examples known to us, with the previous minimum being a single eupathidium, p′, in the enarthronote family Psammochthonidae (Fuangarworn and Norton 2013). The Botswana species of Aquanothrus has both proral setae of tarsus I formed as eupathidia in all instars.
Gnathosoma — Engelbrecht (1975) did not indicate the orientation of the illustrated chelicera (his Fig. 26) but it is abaxial. Seta chb is low on the abaxial face, with Trägårdh's organ (unlabeled) on the opposite side. Seta cha is high on the adaxial face, not the abaxial face, and the movable digit is well inserted into the cheliceral body in the usual manner, so that its base should have been drawn with a broken line. As in Paraquanothrus species, there is a long, looping portion of trachea, terminating in each chelicera. The palp does not vary noticeably among our specimens from Vegkop, Thaba Patchoa and Botswana, but all differ from that illustrated in Engelbrecht's Fig. 27 in several ways (cf. our Fig. 18E, F): (1) tibial seta l′ is present, high on the adaxial side, so the segment has the usual three setae; (2) seta cm is the largest, most proximal seta of the tarsus and dorsally positioned, not a small spine as in his figure; (3) solenidion ω is baculiform, inserted on the abaxial face of the large basal tubercle. It seems clear that Engelbrecht confused seta cm with the smaller ω, but in none of our specimens is any seta as minute and spiniform as that labeled ‘cm’ in his figure. Engelbrecht did not mention the presence of eupathidia, but the same four are present in all studied specimens: acm, (ul) and su. The somewhat separate tubercle of acm is difficult to see in abaxial view, but it is present in all specimens (our Fig. 18F).
Prodorsal setae of larva — Engelbrecht (1975) described and illustrated (his Figs 9, 30) A. montanus as lacking prodorsal setae le and ro. In our specimens from Vegtop, pair ro are present and relatively conspicuous. Pair le are also present, in a position similar to that illustrated for nymphs (his Figs 31, 33, 35), but are small, fine and difficult to see.
Gastronotic sclerites of larva — Engelbrecht (1975) described and illustrated (his Fig. 29) the presence of ‘slightly chitinized sclerites’ on the larval gastronotum (‘notogaster’); two are illustrated, but three are indicated in the text, with the third perhaps being the prodorsum. We studied several larvae from Vegkop Mountain, the type locality, and found no such discrete sclerites. All juveniles have patterns of plication that result in a vague ridge on each side, running longitudinally between setal rows d and l, and in the larva these outline the regions of the supposed sclerites. Perhaps some residual retention of cerotegument pigmentation in this region, when specimens were fresher, gave an impression of a cuticular difference, but there is no sclerite or other formation resembling the porose sclerites of juvenile Podacaridae (Ermilov et al. 2012), or even the leathery patches of Paraquanothrus species.
Claparède's organ — Engelbrecht (1975) did not illustrate (in Fig. 30) or mention this larval organ but it is present. As in Paraquanothrus species it is represented by a porose dome sunken within a cavity. There is no scale-like protective seta (1c), and the seta he labeled as 1c lies just medial to the organ, so is instead probably 1b; 1c appears in the protonymph as in Paraquanothrus.
Paraproctal atrichosy — Engelbrecht (1975) did not recognize that the paraproctal segment is glabrous (atrichous) in the larva, protonymph and deutonymph of A. montanus, as it is in Paraquanothrus species and a majority of other Brachypylina (Grandjean 1949a). Setae of the respective segment are added in the subsequent instar. In his illustrations of the larva (his Figs 29, 30) the setae labeled h2, ps2 and ps3 are respectively h1, h2 and h3. The small, faint structure labeled ps1? in Fig. 30 probably is cupule ip; there is no seta at that location.
Line of dehiscence in juveniles — Engelbrecht (1975) did not illustrate it, but the larva and nymphs of A. montanus and other species have an inconspicuous, permanent line δ. It is U-shaped, incomplete anteriorly, and passes well dorsal to gland opening gla; it effaces at a level just anterior to the periglandular leathery patch. Posteriorly, the line has a normal path, passing above cupule ip in the larva, but below ip in nymphs.
With traits of Aquanothrinae (Engelbrecht 1975 and below). Adult. Cuticle usually with thick, continuous crust of epibiotic diatoms or other organic material (Fig. 18G); invaginated portion of apobasic notogastral seta insertion in form of narrow tube (Fig. 18D). Bothridium on prominent, dark tubercle, bothridial seta attenuate; seta ex distinct, longer than le; in present or absent. Dorso- and pleurophragmata large, conspicuous (Fig. 18A). Dorsosejugal scissure incomplete medially, margin of notogaster projecting anteriorly, merging with prodorsum (Fig. 16E). Lenticulus relatively large, width half mutual distance of setal pair c1 or greater. Notogaster with 10-15 pairs of setae; c2, c3 present or absent. Usually one pair of anal setae, inserted in anterior half of plate. Preanal plate trapezoidal, not wider than long, clearly narrower than vertical wall (Fig. 18B, C). Only coxisternal apodeme apsj perforated (others solid). Ovipositor with three pairs of small coronal setae (k); distal setae of terminal lobes (τ1 on each paired lobe, pair of ψ1 on unpaired ventral lobe) about as long as lobe, extending well beyond its tip. Spermatopositor with six pairs of setae (ψ2 present). Labiogenal articulation complete or incomplete; labrum narrowed distally, with terminal row of cilia. Palp dorsally swollen at base, but only acm on separate, distinct tubercle (Fig. 18E, F); setae acm, su and pair (ul) all eupathidial. Postpalpal seta ep attenuate. Leg pretarsi heterotridactylous. Trochanters III, IV of normal form, with attachment stalk at proximal end of segment. Trochanters III, IV with single trachea and all femora with two tracheae from same stigma; stigmata all high on adaxial face, some nearly dorsal. Famulus minute, vestigial or absent from tarsus I; solenidion ω2 present on tarsus II, adjacent to ω1 in separate alveolus; proral setae of tarsus I eupathidial or not; iteral setae absent from all tarsi; tarsi I, II, IV with paired primiventral setae; pv′ of tarsi I and II unusually high on adaxial face. Genua I, II with large seta d coupled to minute solenidion σ. Seta v′ present on all trochanters. Juveniles. Patch of leathery cuticle only around opisthonotal gland opening. Line of dehiscence (δ) passing above cupule ip in larva, but below ip in nymphs. Protonymphal leg IV setal formula 0-0-0-0-7 (pv″ present). Eggs with dense layer of large tubercles, ca. 5 μm diameter.
Paraquanothrus and Aquanothrus appear to represent sister-groups that are respectively distributed in North America and Africa. Paraquanothrus can be distinguished from Aquanothrus by the following synapomorphies. (1) The inconspicuous opening of the bothridium, flush to the surface (vs. the plesiomorphic elevation of the bothridial wall from the surrounding cuticle). (2) The perforation of coxisternal apodemes ap2, ap3 (vs. solid). (3) The absence (loss) of coronal setae on the ovipositor (vs. presence of normal three pairs). (4) The absence (loss) of distal cilia on the labrum (vs. their presence). (5) The expression of palp tarsal setae acm and su as normal setae, i.e. their eupathidial nature lost (vs. with eupathidial form). (6) The proximal dorsal enlargement of the palp tarsus to incorporate insertions of cm, acm and ω (vs. slight enlargement, with acm maintaining distinct basal tubercle). (7) The unusual structure of trochanters III and IV, with the articulating stalk attached subterminally (vs. typical terminal right-angle bend at proximal end of trochanter). (8) The unusual form of femora III, IV somewhat rectangular in lateral view: proximally broad but abruptly narrowed to articulating stalk, forming vertical face (vs. gradually tapered proximally). (9) The different location of respiratory stigma on femora III, IV (on or near proximal vertical face) compared to that of I, II (low on paraxial face near mid-length of segment) (vs. location similar on all femora). (10) The regressive loss of solenidion ω2 from tarsus II (vs. its presence). (11) Pretarsal monodactyly (vs. tridactyly). (12) The presence of lateral, aggenital and posteroventral leathery patches on the gastronotum of juveniles (vs. their absence). (13) The passing of dehiscence line δ above cupule ip in nymphs (vs. below ip). (14) Several regressive setal losses (vs. their presence): (a) loss of v′ from trochanters I, II; (b) loss of v′ from genua I, II; (c) losses of primiventral setae, including pv′ from tarsus I, pair (pv) from tarsus II, pv′ from tarsus III, and pv″ from tarsus IV.
Aquanothrus can be distinguished from Paraquanothrus by the following synapomorphies. (1) Dorsosejugal scissure medially interrupted, i.e. with medial fusion of notogaster and prodorsum (vs. completely separated). (2) Dorso- and pleurophragmata large, conspicuous (vs. small, inconspicuous). (3) Respiratory stigma on all leg femora nearly dorsal (vs. that of I, II in middle or low position on adaxial face). The coating of epibiotic diatoms or other organic material also seems unique to Aquanothrus. We have seen no diatoms on the cuticle of Paraquanothrus grahami, and those occasionally seen on P. spooneri seem of a normal, free-living type.
If these relationships are true, several incongruencies are apparent. (1) Most obvious is the presence in P. spooneri of saccules on leg femora and porose areas on trochanters III, IV, whereas in P. grahami and species of Aquanothrus all femora and trochanters III, IV have tracheae. As invaginated respiratory surfaces, tracheae have been considered apomorphic relative to saccules (Norton and Alberti 1997). (2) Paraquanothrus spooneri develops iteral setae on legs I-III, but they are lost (apomorphic) from P. grahami and known Aquanothrus species. (3) Paraquanothrus spooneri has unusually elongated tectal setae (tc), an apomorphy shared with A. montanus, but not the other (unnamed) Aquanothrus species.
The close relationship of Paraquanothrus and Aquanothrus seems clear, and classification of the latter in Ameronothroidea and Ameronothridae has been mostly without controversy. But when examined in detail, support for the current concept of both superfamily and family is relatively weak. As recently discussed by Pfingstl (2017), ideas about the composition of Ameronothroidea have varied significantly over time and among authors. When first recognized as a superfamily (Balogh 1961) it included four families: Ameronothridae, Podacaridae, Cymbaeremaeidae and Micreremidae. A decade later (Balogh 1972), Selenoribatidae and Fortuyniidae had been added, with Cymbaeremaeidae and Micreremidae removed to Cymbaeremaeoidea. Weigmann and Schulte (1977) recommended the merger of Podacaridae and the short-lived Aquanothridae with Ameronothridae, a proposal followed by most subsequent authors. Fujikawa (1991) was the most notable exception, maintaining Podacaridae and Aquanothridae, and adding Adhaesozetidae to Ameronothroidea. Behan-Pelletier (1997) then included Tegeocranellidae in Ameronothroidea, as its closest relatives seem to be Selenoribatidae and Fortuyniidae. Norton and Behan-Pelletier (2009) attempted to diagnose the four-family concept—Ameronothridae, Selenoribatidae, Fortuyniidae, Tegeocranellidae—but most listed traits are plesiomorphic or variable (present or absent) and none is an unequivocal synapomorphy of the group.
Concepts of Ameronothridae also have changed, as shown by examples in Table 4. Twelve genera (plus several synonyms) have been included in the family since Vitzthum (1943) first proposed it. We believe that four can be excluded from any modern concept of the family. (1) Selenoribates is the type genus of the well-characterized Selenoribatidae (Pfingstl and Schuster 2012). (2) Adhaesozetes is the type genus of Adhaesozetidae, which has been moved among several superfamilies but seems best accommodated in Licneremaeoidea (Walter and Behan-Pelletier 1993). (3) Seteremaeus was not assigned to a family when first proposed but Hammer (1971) considered it similar to Scapheremaeus. Since then, all but Fujikawa (1991) have included the genus in Cymbaeremaeidae (Cymbaeremaeoidea) (Balogh and Balogh 1992; Subías 2004; Colloff 2010. (4) The case of Capillibates is more complex.
When proposing Capillibates, Hammer (1966) did not discuss family affiliation, but she compared it to two cymbaeremaeid genera, Bulleremaeus and Glanderemaeus, and it was included subsequently in Cymbaeremaeidae by Luxton (1985) and Fujikawa (1991). Without discussion, several other authors (Balogh and Balogh 1992; Colloff and Halliday 1998; Subías 2004) included Capillibates in Ameronothridae, but we see no evidence to support this transfer. The only named species—Capillibates stagaardi Hammer, 1966—is unlike other Ameronothridae in being minute (230 μm), with small anal and genital plates, and having well-developed pedotecta. Few specimens have been collected, but the known habitats are aligned with those of Cymbaeremaeidae. The type series includes two specimens from liverworts and moss in New Zealand, though two others were collected from New Zealand pine-forest soil (Minor 2011). An unnamed species of Capillibates was reported from various arboreal microhabitats in Australia (Walter 1995). Woas (2002) included Capillibates in Micreremidae, another family that includes small arboreal species, but few supporting details were offered for this interesting proposal.
Chudalupia was unknown when Weigmann and Schulte (1977) proposed their broad concept of Ameronothridae, but Wallwork (1981) added Chudalupia without discussing other options. No author has yet questioned its inclusion in Ameronothridae, but despite having a similar habitat, the single known species (C. meridionalis) possesses none of the synapomorphies of Aquanothrinae (see below).


The distribution of adult and juvenile traits of the seven remaining genera in Table 4 was examined with cladistic logic in Weigmann and Schulte's (1977) influential study. They concluded that the gradations and mosaic distributions of characters among the genera could not logically support a separation of Ameronothridae (Ameronothrus), Aquanothridae (Aquanothrus) and Podacaridae (Podacarus, Halozetes, Alaskozetes, Antarcticola, Pseudantarcticola). They found no synapomorphy that could distinguish a separate family Podacaridae (but see Remark 19) and no clear character gaps sufficient to justify recognition of Aquanothridae, though errors and omissions in the original description of A. montanus (see above) partly compromised their dataset. But neither could they propose feasible synapomorphies to support their expanded concept of Ameronothridae (including Ameronothrus, Aquanothrus and the ‘podacarid genera’: hereon called Ameronothridae sensu W-S). The single trait they proposed—the sharing of a plicate (pleated) cuticle in juveniles—we feel is instead a symplesiomorphy (see Remark 2).
Two other studies based on cladistic principles—Behan-Pelletier (1988, 1997)—have addressed Ameronothridae, touching also on Cymbaeremaeoidea and Licneremaeoidea (see also Walter and Behan-Pelletier 1993). Pfingstl and Schuster (2012) compared traits of ameronothroid families, but not with a cladistic approach. Of these studies, only Weigmann and Schulte (1977) analysed Aquanothrus separately, and none provided evidence for a monophyletic concept of Ameronothridae.
Several molecular studies, collectively using a variety of genes, suggest relationships of taxa within Ameronothroidea and Ameronothridae that have little correspondence with current, morphology-based classification, in effect shuffling them with taxa in Cymbaeremaeoidea and Licneremaeoidea. Schäffer et al. (2010), using fragments of 28s rDNA, ef1α, and hsp82 genes, proposed a tree in which none of these three superfamilies is monophyletic. The cymbaeremaeoid sister-groups Cymbaeremaeus and Ametroproctus are distant from the supposedly confamilial Scapheremaeus, with the latter instead nesting among several genera of the licneremaeoid family Scutoverticidae. Ameronothrus also nested within Scutoverticidae, well distant from Podacaridae (Podacarus). The more focused tree of Mortimer et al. (2011), using combined data from the mitochondrial cytochrome oxidase (subunit I) gene and the nuclear histone-3 gene, also indicates the polyphyly of Ameronothroidea: Aquanothrus montanus is the most distant of three outgroups to a monophyletic selection of numerous Podacaridae species. The tree of Krause et al. (2016), based on the 18s rDNA gene, shows Paraquanothrus grahami (listed as ‘Ameronothridae n. gen. near Aquanothrus’) as the sister-group of Scapheremaeus. The Scapheremaeus-Paraquanothrus branch is itself distant from Cymbaeremaeus, and instead is the sister-group of Eremaeozetes (Eremaeozetidae), with Scutovertex as the closest outgroup. The ameronothroid families Selenoribatidae and Fortuyniidae were distant from Paraquanothrus so, effectively, their tree also suggests that none of three superfamilies is monophyletic. Cymbaeremaeus and Scapheremaeus are also widely separated in the trees of Maraun et al. (2009), Iseki and Karasawa (2014) and Xue et al. (2017), all based on the 18s rDNA gene.
The molecular studies seem consistent on three points that are directly or indirectly relevant to Aquanothrus, Paraquanothrus, and Ameronothrus. (1) None of the three genera has a close relationship with Podacaridae. Nor are they closely related to the clade including Fortuyniidae, Selenoribatidae and Tegeocranellidae [these three families form a well-supported monophyletic group (Behan-Pelletier 1997) that we refer to collectively below as the ‘fortuynioid’ families, but imply no formal taxonomic name or rank]. (2) Paraquanothrus and Ameronothrus are close to Scapheremaeus. (3) Scapheremaeus is in turn close to (or within) the licneremaeoid family Scutoverticidae and distant from the supposedly confamilal Cymbaeremaeus and Ametroproctus. The third point is not new: Balogh and Mahunka (1981), and more explicitly Woas (1991, 2002), included Scapheremaeus in an eponymous family, distinct from Cymbaeremaeidae, though it was not named according to international rules. No individual molecular study has included more than one of the three ameronothrid genera, so there is only indirect support for a linkage of Ameronothrus with Aquanothrus and Paraquanothrus, through their mutual relationship to Scapheremaeus.
The molecular results paint a picture in which Aquanothrus, Paraquanothrus and Ameronothrus (Ameronothroidea, in part) are closely related to Scapheremaeus (Cymbaeremaeoidea, in part), which in turn is nested within Scutoverticidae (Licneremaeoidea, in part). There is some modest morphological support for these generic relationships, but it is fragmentary and inconsistent (homoplasious) in some cases, so the following discussion is meant to be heuristic, not a formal cladistic analysis.
We identified only one progressive synapomorphy linking Ameronothrus and Paraquanothrus to Scapheremaeus, and it is inconsistent: the presence of an ascleritic lateral band on the adult notogaster (see Remark 9). Two regressive traits—loss of primilateral and proximal accessory leg setae (Remarks 21, 24)—are shared among the three genera, as well as Aquanothrus. Four other characters support a grouping of Aquanothrus, Paraquanothrus and Scapheremaeus that does not include Ameronothrus. Two of these are progressive synapomorphies: the apobasic insertion of notogastral setae (Remark 10) and the insertion of palp tarsal seta acm on a separate tubercle (Remark 18); while neither is present in Ameronothrus, each is shared with some cymbaeremaeoid and licneremaioid taxa. The other two are regressions of iteral and primiventral setae of leg tarsi (Remarks 22, 23). In general, setal losses are weak synapomorphies, since they are prone to convergence; but they can identify shared trends, i.e., susceptibility to loss of setae that typically are present in Brachypylina. We identified no synapomorphies that link Ameronothrus to Scapheremaeus, while excluding Aquanothrus and Paraquanothrus.
Morphological support for the indirect molecular result—that Aquanothrus and Paraquanothrus are closely related to Ameronothrus—is not overwhelming. The shared regression of pedotecta (Remark 6) could be viewed as the strongest supporting synapomorphy, since it is an unusual trait in Brachypylina. Another synapomorphy that excludes Scapheremaeus is the perforation of at least one apodeme (apsj; Remark 7). Several similarities of Paraquanothrus and Ameronothrus (but not Aquanothrus) are interesting, but inconsistent among species: a weak adult procuticle (Remark 1); the rare loss of eupathidial nature in palp setae acm and su (Remark 17); and the rare coupling of tarsus II seta ft′ with solenidion ω1 (Remark 28).
By contrast, two traits oppose a close relationship of Aquanothrus and Paraquanothrus with Ameronothrus. The notogastral setae of Ameronothrus insert normally, rather than being apobasic; apobasic setae seem pervasive in those outgroups of Aquanothrus and Paraquanothrus that are inferable from molecular studies (Scapheremaeus, Scutoverticidae, Eremaeozetidae). Ameronothrus also lack the tubercle supporting palp seta acm when it is eupathidial, found in Aquanothrus and Scapheremaeus, with some indefinite occurrence in Scutoverticidae (Remark 18).
In this confused context, we follow Pfingstl (2017) in recognizing the separate family Podacaridae (diagnosed by Grandjean 1955 and Wallwork 1964), thereby leaving Aquanothrus, Paraquanothrus, Ameronothrus and Chudalupia in Ameronothridae. A new diagnosis of this family concept—which is supported largely by the shared regression of pedotecta—is offered below. An excellent diagnosis of Ameronothrus can be found in Schubart (1975) and a brief one for Chudalupia in Wallwork (1981), but it should be corrected and emended by material in many of the following Remarks. In some respects, Chudalupia seems the most similar of these genera to Scapheremaeus: it has the deeply black, globular bothridial seta characteristic of most Scapheremaeus, and a strong tibia I apophysis, typical of Cymbaeremaeidae and Scutoverticidae but not found in other Ameronothridae. It has the acm-tubercle and apobasic setae but lacks the ascleritic notogastral band and the various leg setal regressions that link Scapheremaeus with Aquanothrus and Paraquanothrus.
We also resurrect Engelbrecht's (1975) family-group, originally proposed as Aquanothridae, but recognize it as the ameronothrid subfamily Aquanothrinae, which includes Aquanothrus and Paraquanothrus. In part, its resurrection highlights a distinct clade within a family that is not otherwise well-supported. But it is justified also by new knowledge of diversity, including our discovery of two North American species of Paraquanothrus and two unnamed African species of Aquanothrus, with probably others of each genus yet to be discovered. In a practical sense, recognition of the taxon will help prevent this small, but interesting group from being overlooked or subsumed when the evolution and systematics of ‘ameronothroid’ mites are discussed.
Engelbrecht's (1975) diagnosis of Aquanothridae was short and based on only A. montanus, the description of which had important errors and omissions, so we provide a new one for the subfamily. In both diagnoses, traits considered synapomorphies in the context of the respective taxon are underlined.
Adult — Brachypylina with simple form, lacking prodorsal lamella, tutorium, notogastral pteromorphs and octotaxic system of porose organs. nderlineCuticle relatively soft to moderately nderlinesclerotized; cerotegument dense, darkly colored and often with even darker inclusions or nderlinenodules, exposing pale procuticle when removed. Rostrum without proximolateral genal incision or taenidium. nderlinePedotecta I, II absent or represented by vestiges; discrete discidium absent, but with cuticular fold between acetabula III, IV. Dorsosejugal articulation complete or medially incomplete. Notogaster with 10-15 pairs of setae; with or without ascleritic lateral band; lenticulus lens-like or absent. Anterior margin of genital valves aligned approximately with apodeme III; genital papillae flattened or not; coxisternal apodemes perforated or not. Ovipositor relatively short, about equal to body depth when fully extended, with or without coronal setae; spermatopositor of normal, short form, or strongly elongated. Subcapitulum with labiogenal articulation complete or incomplete laterally; without mental tectum or axillary saccule; palp seta acm inserted on tubercle or not, acm not coupled with solenidion ω. Legs with porose organs surficial or invaginated; tibia I with (Chudalupia) or without distal tubercle bearing solenidion φ1; subunguinal seta of tarsus I not eupathidial; solenidion ω2 of tarsus II present or absent.
Juveniles (larva and protonymph unknown in Chudalupia) — Gastronotum with plication closely spaced; without dorsal sclerites; line of dehiscence U-shaped or circular. With paraproctal atrichosy in larva, protonymph and deutonymph; ontogeny of genital setae (larva to adult) 1-3-5-6, all at or near medial margin. Palp femur with two setae in all instars. Legs with seta d present and coupled to solenidion on all tibiae and genua I-III in all juveniles. Protonymphal tarsus IV with six or seven setae. Embryonic development (to prelarva) external or intrauterine.
Adult — Ameronothridae with hysterosomal and basal leg setae apobasic, alveolus at bottom of cup-like or tubular invagination. nderlineTrichobothrium unique: bothridium only slightly S-shaped, nderlinestrongly regressed, tubular throughout; bothridial seta well-developed, setiform, longest non-leg nderlineseta. Interlamellar seta small or absent (present in juveniles). Lenticulus well-circumscribed, lens-like, with underlying red pigment in life. nderlineCoxisternal apodeme apsj perforated; ap2 and nderlineap3 perforated or not. Spermatopositor of normal, short form, with five or six pairs of setae. nderlineGenital papillae of similar size, strongly laterally flattened (about three times longer than broad). Palp tarsus with seta acm eupathidial or normal, nderlineinserted on separate distinct tubercle or sharing nderlinelarger basal tubercle with cm and ω. Rutellum lacking an oncophysis. Chelicera penetrated internally by nderlinelong, serpentine trachea with narrow terminal bulb. Legs with invaginated porose organ (saccule or trachea) on all femora; trochanters III, IV with trachea or porose area; tibiae and tarsi without porose organ. Tibia I without distal tubercle bearing solenidion φ1. nderlineIteral nderlinesetae regressive: absent from tarsus IV; present or absent on I-III (except it′ always absent from III). nderlinePrimilateral setae and proximal accessory setae absent from tarsus I; primiventral nderlineseta pv″ absent from tarsus III, pair present or absent from other tarsi; on tarsus I seta ft″ close to solenidion ω1, in same or adjacent alveolus. Seta d lost from genua I-II, lost or retained on genu III; lost from tibiae III, lost or retained on other tibiae.
Juveniles — Prodorsum nderlineposteromedially with well circumscribed, oval to rectangular nderlinehyaline region. Gastronotum with leathery patch of cuticle around opisthonotal gland opening, with or without other patches in lateral and ventral regions. Line of dehiscence U-shaped, not continuing anteriorly beyond mid-length of gastronotum. nderlineLarva with relatively small nderlineClaparède's organ, retracted under tectum; protective scale-like seta 1c not discerned and no nderlinesetiform homologue present. Protonymphal leg IV with six or seven tarsal setae (pv absent or present), other segments glabrous. Embryonic development external: intrauterine prelarvae not observed.
A close relationship of Ameronothridae with Scapheremaeus would pose some taxonomic problems, but it is an idea worth pursuing, and there is also some ecological congruence. Many Scapheremaeus species are arboricolous (Colloff 2009, 2010), and while most Ameronothrus species are marine-intertidal, the earliest derivative member, A. lapponicus Dalenius, 1963, is an arboricole and several species are associated with supralittoral saxicolous lichens (Schubart 1975). Other Scapheremaeus species inhabit mosses that are subject to intermittent desiccation, including those associated with rock pools (Colloff 2010; Norton et al. 2010), which is the habitat of Chudalupia and Aquanothrinae.
Clearly, the current superfamily concepts of Ameronothroidea, Cymbaeremaeoidea and Licneremaeoidea, e.g. those used by Norton and Behan-Pelletier (2009), Schatz et al. (2011), and to a large extent by Subías (2004), need revaluation and correction. From a morphological standpoint, they are difficult, perhaps impossible, to diagnose without listing mostly symplesiomorphies or variable (present or absent) traits; this is a morphological indication that they are not monophyletic, consistent with the molecular data.
From an evolutionary standpoint, these are important groups that are at or near the base of Poronota, a significant radiation of oribatid mites characterized by the presence of paired dermal glands on the notogaster—the octotaxic system of secretory porose organs (Norton and Alberti 1997). Licneremaeoidea are an obviously paraphyletic grouping of taxa that possess these glands while retaining the plesiomorphic plicate juvenile cuticle and lacking certain specialized defensive structures (particularly pteromorphs) in the adult. For more than a half-century we have recognized that gland presence, form and number are inconsistent in the superfamily. This apparent homoplasy has made interpretation of the early evolutionary dynamics of the octotaxic system difficult (Grandjean 1954a; Norton and Alberti 1997), contributing to an apparent ‘mosaic evolution’ of traits in early-derivative Poronota (Woas 1990, 1991; Weigmann 2010).
Eremaeozetidae, Unduloribatidae, and various genera or species of Scutoverticidae and Micreremidae lack the octotaxic system, but each has close relatives possessing all or part of it. We interpret the absences as losses, a more parsimonious view than invoking multiple evolutions of similar systems. Collectively, the molecular studies cited above suggest that Ameronothridae and Scapheremaeus (but not the supposedly confamilial Cymbaeremaeus or Ametroproctus) also can be counted among the many early-derivative Poronota that lost the system. Less consistently, some molecular phylogenies suggest similar gland losses in more distant branches: Podacaridae (Schäffer et al. 2010), the fortuynioid families (Iseki and Karasawa 2014), Limnozetoidea and Tectocepheoidea (Maraun et al. 2009, Dabert et al. 2010, Pachl et al. 2017). Should we ever develop a stable and well-supported superfamily classification that reflects phylogeny, it will be quite different from the current one.
Collectively, the following Remarks have several aims: (1) to highlight selected aspects of adult and juvenile morphology of Aquanothrinae; (2) to compare Aquanothrinae with other Ameronothroidea, Cymbaeremaeoidea and Licneremaeoidea, in light of potential relationships inferable from the molecular studies discussed above; and (3) to clarify or correct statements in the literature that relate to the individual topics, using internal notes marked with superscripts.
Generalizations about the taxa were gleaned from the following references, augmented by studies of specimens in the first author’s collection. For Ameronothroidea these included: Schubart (1975) for Ameronothrus; Wallwork (1981) for Chudalupia (supplemented by studies of topotypes); Grandjean (1955) and Wallwork (1964) for Podacaridae; Grandjean (1966, 1968) and Pfingstl (2013, 2015a) for Selenoribatidae; Hammen (1963) and Pfingstl (2015b) for Fortuyniidae; and Grandjean (1962) and Behan-Pelletier (1997) for Tegocranellidae. The review by Ermilov et al. (2012) of juveniles in Ameronothridae (sensu W-S) was helpful. For Cymbaeremaeoidea, including only Cymbaeremaeidae, principal references were Travé and Fernandez (1986), Behan-Pelletier (1987, 1988), Woas (2000), Colloff (2009, 2010), Pfingstl and Krisper (2011a) and Ermilov et al. (2015). For Licneremaeoidea we referred largely to the following: Travé (1963b), Woas (1998), Ermilov et al. (2008), Pfingstl et al. (2009a, b, 2010a, b) and McCullough and Krisper (2013) for Scutoverticidae; Behan-Pelletier et al. (2005) for Dendroeremaeidae; Kok (1968), Coetzee (1987), Martínez et al. (1995), Fernandez et al. (1997) and Ermilov and Minor (2015a) for Lamellareidae; Pfingstl and Krisper (2011b) for Micreremidae; Walter and Behan-Pelletier (1993) and Behan-Pelletier and Walter (2007) for Adhaesozetidae; Grandjean (1958a) for Charassobatidae Grandjean, 1958; and Woas (1998) and Martínez and Herrero (2006) for Passalozetidae. The licneremaeoid families Eremellidae and Licneremaeidae are mostly ignored: details of their morphology are poorly known, and we consider neither to have potential as a close relative of Aquanothrinae.
1. Cuticle of adult – In comparing Podacarus auberti with Ameronothrus marinus (Banks, 1896), Grandjean (1955) noted that their juveniles share the unusual dark cerotegument overlying a much lighter procuticle (= ‘cuticle’ in his sense), but that the procuticle of adult A. marinus was usually soft and deformable, whereas that of P. auberti was harder. Schubart (1975) described the procuticle of Ameronothrus spp. as leathery, flexible and only faintly pigmented, contrasting greatly with the dark cerotegument when the latter is removed in patches. He also described the metallic sheen of living mites in reflected light and the lightening of cerotegument in preserved specimens. These descriptors of Ameronothrus also apply to Paraquanothrus grahami. The ultrastructure of cerotegument also is similar in Ameronothrus and P. grahami: cf. Pugh et al. (1987) with Norton et al. (1996b) and our Fig. 3. Its chambered nature may play a role in plastron formation (Messner and Adis 1992, Pfingstl and Krisper 2014) but also may be important when water is absent, by maintaining a buffering layer of humid air, as suggested by Iordansky and Stein-Margolina (1993) for juveniles of xerophilic terrestrial taxa. Collectively, these cuticular traits might suggest a close relationship between Ameronothrus and Aquanothrinae, but precise definitions of characters and states do not yet exist, nor do comparable data for most other groups. Also, the cuticle of P. spooneri appears somewhat harder (more like that of Podacaridae), as suggested by the development of a notogastral tectum.
2. Gastronotic cuticle of juveniles – A variety of cuticular types can be found in juveniles of Ameronothroidea, Cymbaeremaeoidea and Licneremaeoidea, but all are essentially ‘plicate’—having closely to widely spaced pleats on the hysterosoma—which we believe is the ancestral state in Brachypylina (Norton and Behan-Pelletier 1986, 2009). The probable immediate outgroup, the nothrine family Hermanniidae, exhibits this trait strongly, and juveniles of early-derivative brachypyline families such as Neoliodidae, Zetorchestidae and Eremaeidae are plicate. Schäffer et al. (2010), agreeing with Weigmann and Schulte (1977), thought plicate cuticle appeared later in brachypyline evolution, but we believe their conclusion is due to inappropriate character coding. They attempted to test the validity of F. Grandjean’s (1954a) classification of brachypyline families based on juvenile morphology, and failed to consider that plicate cuticle and the carrying of exuvial scalps (eupheredermy) can coexist, as in juveniles of the two basal eupheredermous taxa included in their study: Platyliodes and Zetorchestes.
Various interruptions of otherwise uniform plication can exist, and our knowledge of their nature is insufficient for proposing homologies or character transformations. Among Cymbaeremaeoidea, well-defined paired patches of cuticle that are porose and apparently sclerotized occur in the aggenital, posteroventral and periglandular regions in Cymbaeremaeus and Ametroproctus, but not in Scapheremaeus. Licneremaeoidea are rather diverse. Plication is unbroken and relatively uniform in Scutoverticidae and Passalozetidae and unbroken, but more irregular and less defined, in Charassobatidae. Adhaesozetidae and Micreremidae are distinctly plicate, with aggenital and periglandular porose areas, as well as (except for Adhaesozetes) posteroventral porose areas. Dendroeremaeidae are only weakly plicate, with aggenital, posteroventral and periglandular porose sclerites, as well as some weakly-defined dorsal regions of porosity. In Ameronothroidea, simple, uninterrupted plication occurs in Ameronothrus and Chudalupia, along with a few Podacaridae. Most Podacaridae have plications interrupted dorsally by large porose sclerites (usually paired), with or without aggenital, posteroventral and periglandular sclerites. Instead of porose sclerites, Aquanothrinae have patches of cuticle that appear ‘leathery' rather than sclerotized (though underlying porosity is visible in P. spooneri) and have less distinct plication than the general cuticle: all have periglandular patches, but Paraquanothrus species also have aggenital, posteroventral and unique, elongated lateral patches. In the fortuynioid families, plicae form well-spaced folds, with large, smooth areas that possibly are lightly sclerotized.
3. Line of dehiscence – The line of dehiscence (δ) is a predetermined line of weakness along which the juvenile cuticle splits at the time of molting (Grandjean 1947b; Norton and Kethley 1994) and several of its traits may prove valuable when known in a wider variety of taxa. First, δ may be indicated on the intact juvenile (permanent) or revealed only during molting. It is permanent in Ameronothridae (including Aquanothrus and Chudalupia, new observations), Podacaridae (described only for Podacarus) and Selenoribatidae, as well as in Cymbaeremaeoidea. It is permanent in the licneremaeoid families Adhaesozetidae and Dendroeremaeidae, but we have data for no other groups.
A second trait is whether δ is complete (forming a circular line) or incomplete anteriorly (U-shaped). It is complete in Ameronothrus and Podacaridae (at least in Podacarus), but incomplete in Paraquanothrus, Aquanothrus, Chudalupia and Selenoribatidae. It also varies among Cymbaeremaeoidea, being complete in Cymbaeremaeus but incomplete in Scapheremaeus. For Licneremaeoidea we found data only for Adhaesozetidae and Dendroeremaeidae: the line extends far anteriorly on each side, but an anterior connection was not noted, even though a scalp can become detached in Dendroeremaeidae (Behan-Pelletier et al. 2005). It can be a difficult trait to characterize in the absence of a permanent, well-defined δ, and another complication is that the trait is known to vary with instar in some species (Grandjean 1965a).
A third trait of δ is its path, relative to other cuticular markers, particularly in the posteroventral region where segmental addition occurs. A permanent line δ always runs dorsal to ip in the larva, but in maintaining its relative position it usually comes to lie below ip in nymphs. Among Ameronothroidea, δ has this usual ontogeny in Aquanothrus and Chudalupia, but δ remains dorsal to ip in all juveniles in Paraquanothrus, Ameronothrus and Podacaridae (from Podacarus). There are few data for the other superfamilies: in Cymbaeremaeoidea, δ shifts below ip in nymphs of Scapheremaeus, but we have found no data for Licneremaeoidea.
4. Notogastral lenticulus and prodorsal hyaline region – A secondary light receptor, in the form of a notogastral lenticulus, is present in adults of some taxa in each of the three superfamilies, but absent from others, and the absences probably represent multiple losses (Weigmann and Schulte 1977). Among Ameronothroidea a lenticulus is present in all taxa except Selenoribatidae and Ameronothrus a. Among Cymbaeremaeoidea, only Scapheremaeus have a lenticulus. Among Licneremaeoidea it is absent from Adhaesozetidae, Lamellareidae and Charassobatidae, but present in Passalozetidae, Licneremaeidae, Dendroeremaeidae, and some genera of Scutoverticidae (Scutovertex, Ethiovertex) but not others. Paired, red pigment-cells are clearly present under the lenticulus in adult Paraquanothrus (Fig. 16A; Supplemental video), and in life they seem similar to those of the well-studied genus Hydrozetes (Alberti and Coons 1999). Red pigment also seems present under the lenticulus of Fortuyniidae (Pfingstl and Krisper 2014, their Fig. 2), but in most cases we have no data from living mites, and the pigment does not persist after preservation.
We suspect that the distinct and well-circumscribed hyaline area at the base of the prodorsum in juvenile Aquanothrus and Paraquanothrus also is associated with reception of ambient light. Red to orange color can be seen by transparency in living juvenile P. grahami (Supplemental video), but we observed no paired pigment-cells like those of the adult lenticulus. Possibly the color derives instead from pigmented body contents, presumably derived from Philodina rotifers, but this would not preclude a role in light reception. Pigment-cells are not required for light sensitivity, as noted by Alberti and Fernandez (1990); they found adults of Oribatella and Chamobates species to have light-sensitive lamellated bodies, even though the notogastral photoreceptor (‘clear spot’) lacks pigment-cells. Ultrastructure studies might similarly reveal lamellated bodies under the hyaline region of juvenile Aquanothrinae. Because of its posterior, central position on the prodorsum, the hyaline cuticle may allow light to impinge directly on the brain and to affect physiology or behavior, even though light probably must pass through the cheliceral retractor muscles that have their origin in this area. Regardless of its function, we consider the prodorsal hyaline region in juveniles to be an autapomorphy of the subfamily. A prodorsal hyaline region is found in adults of the macropyline genus Galapagacarus (Hermanniidae; Alberti and Moreno-Twose 2012), but it has not been closely examined and juveniles are unknown. Several other literature indications need explanation. Wallwork (1981, his Fig. 5) illustrated a rectangular area at the base of the prodorsum in a tritonymph of Chudalupia meridionalis, but it is only a well-circumscribed topographic area of non-porous cuticle, not a hyaline region, and it is covered with dark, granular cerotegument indistinguishable from that covering the rest of the prodorsum. Schäffer and Krisper (2007) considered a small area on the prodorsum of nymphal Scutovertex minutus (Koch, 1836) to be a lenticulus, inferring it to be homologous to the notogastral lenticulus of the adult; however, its properties were not described, nor was its novel position discussed. McCullough and Krisper 2013 made no mention of such a ‘lenticulus’ on the prodorsum of S. pannonicus Schuster, 1958 juveniles, but they did discuss a possible prodorsal origin for the ‘corneal’ cuticle of the lenticulus of the adult. We examined juveniles of two unidentified species of Scutovertex, and they have no prodorsal hyaline region.
a Alberti and Coons (1999; their Fig. 243B) included a label for the lenticulus on a dorsal figure of Ameronothrus, but this was an error: the structure is simply the medial region of the notogaster that encroaches on and fuses with the prodorsum.
5. Dorso- and pleurophragmata – Most muscles that operate the chelicerae and subcapitulum originate on the prodorsum, where their sigilla are often visible by transparency and described as ‘maculae' or ‘spots' in the sclerotized adult. This is the state in early-derivative Brachypylina and many middle-derivative groups, but in many adult Brachypylina these muscle attachments (as well as some dorsoventral muscles of the hysterosoma) attach instead to discrete apodemes that extend internally from the posterior edge of the prodorsum. The apodemes vary in form and number, but exist most often as an unpaired, or closely paired dorsophragma (= opisthophragmatic process or apophysis) on which cheliceral muscles originate, and a paired, often larger pleurophragma in the post-bothridial region on which other cheliceral and some subcapitular muscles originate. Typically, as in Paraquanothrus, these gnathosomal muscles attach to the ventral face of the apodeme while hysterosomal muscles attach dorsally. Phragmata often are unmentioned or poorly represented in descriptions, so generalizations about their distribution are tenuous. Among Ameronothroidea, phragmata are absent from Ameronothrus, but present (weakly to strongly developed) in Chudalupia, Aquanothrinae, Podacaridae, Tegeocranellidae and Fortuyniidae. Discrete phragmata have not been described for Selenoribatidae, but there is an apodematic band (Grandjean 1968) with unknown association to musculature. Some Cymbaeremaeoidea have distinct phragmata (Scapheremaeus) but these structures have not been reported in others (Cymbaeremaeus, Ametroproctus). Among Licneremaeoidea, they are present in Scutoverticidae, Dendroeremaeidae, Lamellareidae and Charassobatidae; they are reported to be absent from Micreremidae (Behan-Pelletier 1997) and are either present (Phylleremus) or apparently absent (Adhaesozetes) in Adhaesozetidae. Homoplasy is indicated by the presence of both states in each of the three superfamilies, though some apparent absences (e.g. Selenoribatidae, Micreremidae) may result from modification or loss.
6. Pedotecta – Scale-like tecta that overreach and protect the respective acetabulum (I, II) from behind (Grandjean 1952) are plesiomorphically absent from early-derivative groups (Hermannielloidea, Neoliodoidea), but they exist in most Brachypylina. Pedotectum I and (usually) II consistently exist in Licneremaeoidea and Cymbaeremaeoidea, but vary within Ameronothroidea. They exist in Podacaridae and the fortuynioid families (II is absent from Selenoribatidae and some Tegeocranellidae), but are essentially absent from Ameronothridae. Weigmann and Schulte (1977) interpreted their absence as secondary (a loss) and—considering the relatively derived position of these groups in molecular studies—we agree with their interpretation. We interpret the vertical ridge posterior to acetabulum I and the epimeral tooth and carina posterior to II in Paraquanothrus grahami as vestiges of pedotecta I and II respectively. These interpretations are tentative: e.g., a ridge somewhat similar to vr exists in Podacarus, despite the presence of a normal pedotectum I (Grandjean 1955). A highly reduced pedotectum I is present in one species of Ameronothrus (Schubart 1975), causing Weigmann and Schulte (1977) to dismiss the absence of pedotecta as a synapomorphy of Aquanothrus and Ameronothrus, but we view it differently: strong pedotectal diminution is a synapomorphy of Ameronothridae, and the total loss of pedotecta is a continuation of the trend in all or some species of each genus.
Grandjean (1947a) had contrasting ideas, believing that the absence of pedotecta (‘lames tectopediales’) in Ameronothrus is plesiomorphic (‘une moindre évolution’). It was one of several traits that led him to envision Ameronothrus as the most primitive genus among that large group of oribatid mites with a ‘normal’ tracheal system, which includes most of the Brachypylina.
7. Coxisternal apodemes – Typically in Brachypylina, four pairs of coxisternal apodemes project internally from the ventral plate to serve as attachment locations for muscles: ap1, ap2, apsj and ap3. They are frequently described incorrectly in the literature, as many authors have confused apodemes with epimeral borders, which are more conspicuous and partly confluent with apodemes when viewed ventrally in transmitted light (Grandjean 1952). The form of apodemes is rarely described, as it requires dissection and viewing in an anterior or posterior direction, so there are relatively few data. The plesiomorphic state for the groups in question is a flat, solid cuticular plate, as found in many early- to middle-derivative Brachypylina—e.g. Eremaeidae (Grandjean 1963a), Damaeidae (Grandjean 1960) and Damaeolidae (Grandjean 1965b). A derived trait is for a particular apodeme to be perforated (troué, Grandjean 1952). Perforation of ap2 can be so extensive that the apodeme has the form of an arch, as is typical of Oripodoidea (e.g. Grandjean 1956b, 1963b).
In Aquanothrinae, at least apsj is perforated. In Aquanothrus (only the Botswana species could be studied) ap1, ap2 and ap3 are solid, while apsj has a small, narrow perforation. In both species of Paraquanothrus, ap1 is solid but the other three are perforated (Fig. 4D). Regarding other Ameronothroidea, we could find only three references to this trait in the literature. Grandjean (1955, 1962) noted that all apodemes were solid in Podacarus and Tegeocranellus. Schubart (1975) found apsj and ap3 to be perforated in Ameronothrus; most species had a solid ap2, but he thought it was possibly perforated in A. lapponicus, the most plesiomorphic of known species.
Considering this shortage of information, we made the following new observations. An undescribed species of Ameronothrus from California has the posterior three apodemes perforated. Apodemes are solid in the single species of Selenoribatidae (Thalassozetes sp.) and Fortuyniidae (F. elamellata Luxton, 1967) that we examined. They share with Tegeocranellidae (T. muscorum Behan-Pelletier, 1997) an unusual state of ap3—the pair meet at the midline to form a single, thin, ligament-like curtain of uniform height; this should be further examined as a possible synapomorphy of the fortuynioid families (see Behan-Pelletier 1997). Chudalupia has solid apodemes. Among Cymbaeremaeoidea, no species seems to have been studied in this regard: we examined Cymbaeremaeus cymba (Nicolet, 1855) (which lacks ap3), Ametroproctus oresbios Higgins and Woolley, 1968, and an unidentified species of Scapheremaeus (from Oregon), and all apodemes are solid. Among Licneremaeoidea the only published information concerns Provertex delamarei Travé, 1963 (Scutoverticidae), which according to Travé (1963b) has the posterior three apodemes perforated; however, we found all apodemes to be solid in Scutovertex cf. sculptus Michael, 1879 (specimens from New York).
8. Genital papillae – Genital papillae are eversible osmoregulatory structures normally retracted into the walls of the genital vestibule (Alberti 1979). They are plesiomorphically round in cross-section, or slightly elliptical, but they have become modified in various oribatid mite taxa (Behan-Pelletier 1991), including size and form changes, and specialization of the anterior papilla. Among Ameronothroidea all three pairs are similar, but they can collectively differ in form among taxa: in Aquanothrinae, Chudalupia and some Podacaridae (Podacarus, Alaskozetes) they are strongly laterally compressed, such that their cross-sectional width is only about one-third its length. In other ameronothroids they are not extremely flattened (at most twice as long as wide), but may be strongly modified in other ways (e.g. Grandjean 1968). We know of no examples of extreme flattening in Cymbaeremaeoidea or Licneremaeoidea.
9. Ascleritic band on notogaster – As discussed by Woas (1991), Ameronothrus and Scapheremaeus share a partial or complete, narrow, submarginal band of soft, desclerotized cuticle on the notogaster (‘circumferential scissure’ of Colloff 2010). Schubart (1975) who coined the term ‘ascleritic band’, considered a complete band to be ancestral in Ameronothrus, with partial bands being derived. A partial band (asc) is found in Paraquanothrus spooneri but at most a vague and variable anterior indication exists in P. grahami; Aquanothrus species lack the structure, as does Chudalupia. We are not aware of other occurrences, but the trait has not been well-surveyed.
10. Setal insertions on notogaster – The body setae (as well as some basal leg setae) of adult Aquanothrinae are apobasic. The term was coined by Grandjean (1956a) in reference to Galumnidae, where they are typical, but apobasic setae are equally characteristic of other large groups, most notably Ptyctima. The cuticle at the site of an apobasic seta forms an abrupt depression—ranging from a cup to a deep, narrow tube, open to the environment —at the bottom of which is a normal setal insertion. Internal to the hidden, ‘sunken’ insertion there is the usual simple cuticular canal through which sensory dendrites pass before attaching to the setal root (the short, usually swollen part internal to the attachment of the soft ring of articulating cuticle). External to the hidden insertion the seta passes through the cup or enveloping tube before emerging at the surface. This differs from setae that insert on a solid, sclerotized structure such as a lamella or tectal limb, where the alveolus is normal and at the surface, but the alveolar canal is greatly elongated to reach the living epidermis. Grandjean (1956a) was uncharacteristically vague about the structure of an apobasic seta, stressing that it emerges from the surface at some distance from where the canal seems to be leading; his Fig. 1E shows the form well, but omits the ring of articulating cuticle just external to the swollen root, such that the seta seems to float, unconnected. The definition of Hammen (1980) is misleading: apobasic setae are described as having a long setal ‘root’ that is far removed from the ‘alveolus': in his view, the ‘root’ seems to be the external part of the seta that traverses the open-air tube or cup, with the ‘alveolus’ being the point where the seta emerges at the surface.
The apobasic setal insertion is a derived state. Its full taxonomic distribution is unknown, but, given its presence in groups as disparate as Ptyctima and Galumnidae, it has evolved more than once. Based on literature and new observations the following generalizations can be made about the form of setae on the adult notogaster of the three superfamilies. No mixed forms were encountered: e.g., on a given specimen all notogastral setae were apobasic or all were normally inserted.
Among Ameronothroidea setae are apobasic in Aquanothrinae and Chudalupia; setae are normal in Ameronothrus, Podacaridae and the fortuynioid families. In Cymbaeremaeoidea setae are apobasic in Scapheremaeus but normal in Cymbaeremaeus and Ametroproctus. In Licneremaeoidea, setae are apobasic in Charassobatidae, Micreremidae and Scutoverticidae (Scutovertex, Arthrovertex, Provertex, Exochocepheus examined), but normal in Passalozetidae, Licneremaeidae, Adhaesozetidae, Dendroeremaeidae, Eremellidae and Lamellareidae. Another questionably classified family, Eremaeozetidae (Eremaeozetoidea), also have apobasic notogastral setae; these mites much resemble Scutoverticidae and representatives of the two families cluster with Scapheremaeus and Paraquanothrus in the molecular tree of Krause et al. (2016).
11. Bothridial seta (sensillus) – The typical bothridial seta (= sensillus) of oribatid mites could be described as an apobasic seta (see above), with the depression being complex: a cup-like outer portion and tubular, usually S-shaped inner portion at the bottom of which is the true insertion. In this form, the seta is highly sensitive to slight air movement and seems to be an anemoreceptor (Alberti et al. 1994 and cited papers). In Aquanothrinae the seta is well-developed, but the bothridium is reduced to a narrow tube, restricting deflection, probably reducing sensitivity, and making the analogy to an apobasic seta seem especially appropriate.
Diminution of the trichobothrial nature of this seta—regression of the bothridium, the seta, or both—is scattered in the oribatid mite phylogenetic tree, but the biological contexts of convergences can be quite different. In most Nothrina, for example, regression affects juveniles but not adults. Since microhabitat generally does not change during their development, there is no obvious adaptive value that relates to environment; instead it seems a fixed phylogenetic trait in the group, one that even carries into the derivative, paedomorphic hyporder Astigmata (Norton 1998). In contrast, some regressions seem induced by the lack of relevance of an anemoreceptor in a particular environment. This would include reduction or loss in juveniles of taxa that develop as burrowers within plant or fungal tissue (e.g. Phthiracaridae, Carabodidae, Hermanniellidae); their adults live outside such substrates and have typical bothridial setae.
Taxa that are active under water commonly show some form of trichobothrial regression, as might be predicted in such a viscous medium. Perhaps best known is Limnozetoidea, in which the trichobothrium of adults ranges from well-developed and clavate to highly regressed, according to species, but all juveniles have a fine, thread-like bothridial seta with a small bothridium (Grandjean 1948, 1951; Behan-Pelletier 1989). By contrast, groups that occupy surface films or simply survive temporary inundation usually retain a typical bothridial seta in all instars. Pauly (1956) suggested that regressed trichobothria may be functional under water, perhaps as current-receptors (rheotaxis). In cases where the seta remains well-developed but the bothridium regresses, such as Aquanothrinae, juveniles of Limnozetoidea (op. cit.), and the aquatic macropyline genus Mucronothrus (Norton et al. 1996a), some functionality seems likely. By contrast, Schubart (1975) considered a rheotactic function unlikely in those Ameronothrus species having highly regressed trichobothria, since the minute seta does not project above the cerotegument.
A typical bothridial seta is found in all Cymbaeremaeoidea and Licneremaeoidea, and in the fortuynioid families of Ameronothroidea. The reduction of the bothridium (but not its seta) is an autapomorphy of Aquanothrinae. Most Ameronothrus species retain the typical form, but species that inhabit periodically flooded coastal areas have regressed both the bothridium (to a tubular invagination) and the seta (to a minute thread) or lost them altogether. The taxonomic distribution of such regressions suggests evolutionary convergence within Ameronothrus (Schubart 1975). Similarly, nearly all Podacaridae have a typical bothridial seta but in at least one species of Halozetes it can be absent from the larva (Engelbrecht 1974). The adult of Halozetes necrophagus Wallwork, 1967 has a well-developed bothridial seta but the bothridium (= pseudostigma) is ‘reduced to a small pit’ (Wallwork 1967); only one specimen was found and the figure is not detailed, but of all known ameronothroid species it seems to have a bothridial seta most similar to that of Aquanothrinae. We agree with Weigmann and Schulte (1977) that trichobothrial regression is convergent within Ameronothroidea, but believe their treatment (presence or absence of the sensillus) was overly simplistic.
12. Gastronotic setation of larva – Clearly, a complement of 12 pairs of gastronotic setae in the larva is ancestral among the taxa under consideration, with the missing seta probably being pair f1 (Grandjean 1954a). A derivation is for seta h3 also to be lost (or represented by a vestige) in the larva, but added in the protonymph and retained for the rest of ontogeny. This development seems odd, but it is found in various scattered families of Brachypylina (Grandjean 1949a, Behan-Pelletier 1997).
Cymbaeremaeoidea retain h3 in all instars as do Licneremaeoidea, other than Lamellareidae, which lack it in the larvaa; Adhaesozetes has h3 in the larva but lacks p3 and c3 throughout ontogeny. Among Ameronothroidea h3 is lost from the larva in the fortuynioid families and one member of Aquanothrinae (P. spooneri); by contrast, Ameronothridae, Podacaridae and the other Aquanothrinae retain the plesiomorphic presence of larval h3. It seems likely that the regression of h3 in the fortuynioid families is a synapomorphy of the group (Behan-Pelletier 1997), while that of P. spooneri and Lamellareidae represent separate convergences. A unique derivation has been described for the podacarid subspecies Halozetes marinus devilliersi Engelbrecht, 1974, which apparently lacks la from the larva (as well as the protonymph)b.
a Martínez et al. (1995) noted that the larva of Tenuelamellarea argentinensis Martínez, Velis, Eguaras and Fernández, 1995 forms only two setae in the h-row and adds a third in the protonymph. However, their labeling of the larval setae (their Fig. 4C) seems incorrect, since the anterior and posterior of the two setae are respectively denoted as h1 and h2; these labels should be reversed.
b As described by Engelbrecht (1974), the gastronotic ontogeny of the podacarid subspecies H. marinus devilliersi would be unique among oribatid mites. However, as in his 1975 description of Aquanothrus (see above), he did not recognize the changes associated with paraproctal atrichosy, and therefore misinterpreted some posterior setae of the larva (his Figs 8, 9): his h2, p2, p3 are actually h1, h2, h3 respectively.
13. Gastronotic setation of nymphs – Ancestrally, the three setae of the p-row join the gastronotic region in the protonymph, resulting in a plesiomorphic setation of 15 pairs, as f1 is lost throughout ontogeny—the condition of ‘unideficience’ (Grandjean 1954a). Ameronothroidea have unideficient nymphs, with three exceptions known to us. Paraquanothrus spooneri is unique in lacking p1 and p2 from all nymphs (as well as the adult). By contrast, Chudalupia lacks p3 in the tritonymph (Wallwork 1981, his Fig. 6; no other juvenile instars were described, but it was also absent from the deutonymphs we studied). The third exception is the podacarid subspecies Halozetes marinus devilliersi, for which a strange and unique development of dorsolateral setae in nymphs has been describeda. No Cymbaeremaeoidea known to us has further setal losses in nymphs. Among Licneremaeoidea, Scutoverticidae, Micreremidae, Passalozetidae, Dendroeremaeidae and Charassobatidae are similarly unideficient, but Licneremaeidae, Lamellareidae and Adhaesozetidae lack seta p3 in nymphs, with Adhaesozetes also lacking c3 (in all instars).
a Though not mentioned by Engelbrecht (1974), seta la of H. marinus devilliersi (apparently absent from the larva; see above), does not appear on either side of his protonymph figure (Fig. 10). By contrast, the two dorsolateral setae symmetrically shown on the deutonymph (his Fig. 12) are labeled la and lm, implying the addition of la and the loss of lp between protonymph and deutonymph; he did not discuss the discrepancy. His figure of the tritonymph (Fig. 14) shows asymmetry, with the usual three dorsolateral setae on the left side but only one (lp according to position) on the right. The labeling of la as the more anterior ‘lp’ in his Fig. 14 is clearly a lapsus. From the apparent variation in the tritonymph, we infer that the odd complements of dorsolateral setae in earlier instars may also represent variation, rather than a unique fixed development.
14. Further setal losses on adult notogaster – While the complement of gastronotic setae in juveniles varies little among the three superfamilies, setation of the adult notogaster is made relatively diverse through various losses. The plesiomorphic state of unideficience—lacking only seta f1—is retained from the tritonymph in some members of each of the three superfamilies, while other members lose as many as six additional pairs, beyond those that may already be absent from the nymphs (see above). In most instances, losses form ‘normal’ patterns that are common to many taxa of Brachypylina (Grandjean 1934a, 1949a). A setation of 14 pairs usually indicates the loss of c3, 13 the additional loss of c1 and 10 the still additional loss of dorsocentral setae (da, dm, dp); 12 usually indicates the loss of all c-setae and retention of dorsocentrals. Since losses incurred in the nymphs are never regained in the adult, focusing on additional losses, rather than total losses, can make adult setation an independent character.
Among Ameronothroidea, Ameronothrus and Chudalupia retain the tritonymphal setation (15 and 14 pairs, respectively). The same is true of the aquanothrine genus Paraquanothrus, but Aquanothrus varies: no losses (15 pairs), or unique loss patterns for 12 and 10 pairs. Podacaridae have 10-15, Selenoribatidae 13-15, and Fortuyniidae 14, all following the normal loss patterns. Tegeocranellidae have 10 or 12 pairs, but the complement of 12 is unique, including c2 while lacking da. Among Cymbaeremaeoidea, with some exceptions in the genus Scapheremaeus (see Colloff 2009), adults have 10-15 pairs, in the normal patterns. Scapheremaeus zephyrus Colloff, 2010 has nine pairs (lacking also p3) and S. longilamellatus Mahunka, 1985 appears to have seven pairs (lacking p2, p3 and la), but their tritonymphs are unknown so the timing of the losses is also unknown. Licneremaeoidea have a similar range, except we know none with 15 pairs. Micreremidae lose c3 (14 pairs) and Scutoverticidae have the same setation, at most, but may have as few as 10 in the normal patterns. Charassobatidae have 13 pairs and Passalozetidae and Dendroeremaeidae have 10, all in the normal patterns. Lamellareidae and some Adhaesozetidae (Phylleremus) lose five in the normal pattern, while Adhaesozetes loses none in the adult. Overall, the range of notogastral setations is similar among the three superfamilies, and we agree with Weigmann and Schulte (1977) that notogastral setations show too much regressive convergence to be highly informative as a phylogenetic character for the groups in question.
15. Notations of coxisternal setae – The chaetotaxy used for setal pairs on oribatid mite epimeres was established by Grandjean (1934b) with two different aspects to the assignment of notations. In the absence of neotrichy, setae of a given epimere typically are arranged in a single transverse, oblique, or curving row, and each pair is assigned a two-part notation: a number for the epimere (1-4) and a letter for its position in the row (a, b, c, d ...) beginning at the midline. This positional criterion is used for the larva, but a second, ontogenetic criterion was applied to setae that are added during development: a sequential letter is given to each newly-appearing pair without concern for position.
Epimere I typically does not add setae during ontogeny, but 1c becomes more easily discerned as it changes form, from a scale that protects Claparède’s organ in the larva to a normal seta in the protonymph. If seta 1c is truly absent from the larva, it may appear in the protonymph (e.g., Aquanothrinae). With few exceptions, epimere II retains a single pair, 2a, in all instars. Epimere III typically has two pairs in the larva, with 3a on soft medial cuticle and 3b lateral to it on the epimeral sclerite; 3c, and often 3d, are usually added sequentially and laterally, after the protonymph. For epimeres I-III this chaetotaxy has been applied quite consistently in the literature, but not so for epimere IV, primarily due to an apparent incongruence in setal positions.
In the large majority of oribatid mites, from all major groups, a single pair of setae appear when epimere IV first forms in the protonymph, called 4a because of the ontogenetic criterion. The pair are widely spaced and each seta is inserted on its epimeral sclerite in a position analogous to 3b on epimere III. The second pair to appear are typically deutonymphal and designated 4b using Grandjean’s ontogenetic criterion; the setae insert well medial to 4a, in a position analogous to 3a and usually also in soft cuticle (e.g., Grandjean 1934b, 1954b, 1963c). Pair 4c, if they appear, are lateral, near the trochanter, in a position analogous to 3c. Authors who disregard the ontogenetic criterion of the chaetotaxy, or lack ontogenetic data, often reverse the notations of 4a and 4b, such that their letter designations are sequential from medial to lateral, as on epimere III. The two approaches result in inconsistency in notations for epimere IV setae in the literature. Even numerical formulae can be affected, since the medial, deutonymphal seta of epimere IV can be so close to 3a as to be incorrectly counted among epimere III setae (Grandjean 1934b, Sidorchuk and Norton 2010).
The ontogenetic pattern described above is found throughout Ameronothroidea, Cymbaeremaeoidea and Licneremaeoidea, with very few exceptions. The notations assigned by Engelbrecht (1975) to Aquanothrus montanus, for example, show the proper application of Grandjean’s chaetotaxy. The most striking exception is in Selenoribatidae, where most coxisternal setae have been lost (Grandjean 1966, 1968; Pfingstl and Schuster 2012). Larvae of this family have only setae interpreted as 1b and 3b; the soft medial region is glabrous, and the setae have positions similar to their perceived homologues in more complete setations, on the epimeral sclerites. One pair are added in the protonymph and given the notation 4a, presumably based on the ontogenetic criterion; they have the same, relatively lateral position on the sclerite as does 4a in other families, i.e. similar to that of 3b.
A less striking, but no less interesting exception is found in Paraquanothrus. While the epimeral ontogeny of P. grahami (Fig. 7B, D) is typical and virtually identical to that of A. montanus, that of P. spooneri (Fig. 13B, D) differs in an illuminating way. Epimere III of the larva bears a single, laterally inserted pair that are obvious homologues of 3b in P. grahami; the medial, soft cuticle is glabrous, with 3a absent as in Selenoribatidae. The second pair of epimere III setae are added medially in the protonymph, in precisely the same position as 3a in P. grahami. The simple and inescapable conclusion is that 3a is delayed one instar during the ontogeny of P. spooneri, relative to P. grahami.
Added to the loss (Selenoribatidae) or delay (Aquanothrinae) of seta 1c, these exceptions show that regressions and additions of epimeral setae, either in phylogenetic or ontogenetic time, can be independent of position. If pair 3a can be lost (Selenoribatidae) or delayed (P. spooneri) while 3b is unaffected, then the same should be possible on epimere IV. In our interpretation, epimere IV of P. spooneri has a setal ontogeny identical to that of epimere III, except that it is delayed one instar due to the protonymphal appearance of the epimere: pair 4b are protonymphal, 4a are deutonymphal. Similarly, we would denote the single epimere IV pair in Selenoribatidae, also protonymphal, as 4b.
We think this approach can and should be applied throughout oribatid mites. If we discard the ontogenetic criterion and focus on relative setal positions to identify homologues, as is done in other aspects of oribatid mite chaetotaxy, then the apparent oddity of epimere IV setation—in which 4b are medial to 4a—ceases to exist.
16. Labiogenal articulation – The labiogenal articulation is a narrow band of soft cuticle running from the mouth toward the insertion of the palp trochanter in Brachypylina, which allows relative movement between the subcapitular mentum and the paired genae (the ‘diarthric’ state of Grandjean 1957). In some Ameronothroidea the articulation is incomplete in adults, not reaching the base of the palp. It can be complete or incomplete in Podacaridae, according to genus, and in Ameronothrus, according to species. It also varies among species in Aquanothrinae, being complete in P. spooneri and A. montanus, and incomplete in P. grahami and the undescribed Aquanothrus species from Botswana. It is complete in Chudalupia and the fortuynioid families, and in all Cymbaeremaeoidea and Licneremaeoidea known to us.
Without explanation, Weigmann and Schulte (1977) considered the incomplete state as plesiomorphic, but we believe it is apomorphic, since the articulation is complete in adults of the earliest-derivative brachypyline groups (Hermannielloidea, Neoliodoidea). It is common for juveniles of Brachypylina to have an incomplete articulation, regardless of the adult state, so we interpret the very homoplasious adult distribution as resulting from multiple independent occurrences of paedomorphosis. A relatively weak subcapitular cuticle may allow enough flexing to make the full articulation less important.
17. Complement of eupathidia on adult palp tarsus – Setae are normally solid mechanoreceptors, innervated only at the base, but on the tarsus of the palp and leg I, as well as on the genitalia, some are usually eupathidia: hollow gustatory receptors with chemosensory dendrites exposed at one or more terminal openings (Alberti and Coons 1999). Setae of the palp tarsus that are most consistently eupathidial in adult Brachypylina are the ultimal pair (ul), followed in frequency by the unpaired anteroculminal, acm, and subultimal, su. The former two do not change during ontogeny, but acm (usually) and su (always) begin as normal setae and attain the eupathidial state sometime after the larval instar (Grandjean 1946b). We consider this pattern of four eupathidia to be plesiomorphic among the three superfamilies. It is the only pattern described for Cymbaeremaeoidea and Licneremaeoidea, but data are few and fragmentary since the eupathidial nature of setae often goes unreported. Among Ameronothroidea the plesiomorphic pattern exists in Podacaridae, the fortuynioid families and Chudalupia, but Aquanothrinae and Ameronothrus are mixed. Aquanothrus retains the four eupathidia, but in Paraquanothrus species both acm and su are normal setae in all instars. Ameronothrus seems to show a similar dichotomy, with most species retaining the plesiomorphic pattern but some expressing acm and su only as normal setae (Schubart 1975). We consider the failure of acm and su to become eupathidial as independent derivations within some Aquanothrinae and some Ameronothrus.
18. Insertion of palp seta acm – In macropyline oribatid mites, seta acm inserts like other setae, without special underlying structure. This probably is plesiomorphic in Brachypylina, but in some members of Neoliodoidea acm is on a distinct tubercle, as in e.g. Neoliodes theleproctus (Hermann, 1804), or a conical mound, as in Poroliodes farinosus (Koch, 1839), and a similar range of form is found in Plateremaeoidea (Woas 1992, 2000), which makes the polarity of this trait somewhat equivocal for the three superfamilies in question. The insertion of seta acm on an isolated, distinct tubercle has been considered an apomorphic trait that characterizes Cymbaeremaeidae (Behan-Pelletier 1987, 1988), though it is only weakly developed in some Scapheremaeus species (e.g. Travé and Fernandez 1986). In Licneremaeoidea acm lacks a distinct basal tubercle, but a broad, volcano-shaped swelling exists in the scutoverticid genus Provertex (Travé 1963b) and possibly some Lamellareidae (Kok 1968). Among Ameronothroidea, acm is not isolated on a basal tubercle with two exceptions: Chudalupia has a distinct tubercle, similar to the common form in Scapheremaeus; and Aquanothrus, in which the eupathidial acm sits on a small tubercle that projects from a modest proximo-dorsal swelling. In Paraquanothrus acm is a normal seta, without a basal tubercle, but it inserts along with cm and ω on a proximo-dorsal apophysis. We consider the Paraquanothrus type to be apomorphic within Aquanothrinae, perhaps with the loss of the small basal tubercle being correlated with losing the eupathidial nature of acm.
19. The ‘double-horn' – The double-horn (‘corne double’ of Grandjean 1946b) is a compound sensory organ on the palp tarsus of many Brachypylina. It involves the whole or partial attachment of two chemoreceptive structures that terminate together: solenidion ω and eupathidium acm—thought to be olfactory and gustatory receptors, respectively—with the solenidion always being proximal to the seta. Grandjean (1965b) explained their union as the formation of a compound organ allowing both smell and taste at the same spot, when the palp tarsus touches an object. The first survey of its distribution (Grandjean 1954a) and subsequent work has shown the double-horn to be a nearly ubiquitous trait of Poronota (Scutoverticidae, Zetomotrichidae and Stelechobatidae are exceptions), but it is present also in a few non-poronotic taxa, including Limnozetoidea, Gustaviidae, Peloppiidae (some, see below), Compactozetidae (including Cepheidae auct.) and Tectocepheidae. The latter family have a rather clear association of ω and acm that Grandjean (1954a), for some unknown reason, did not consider a double-horn (cf. Suzuki 1978, his Fig. 1E, which is typical of the genus).
But double-horns are not all equal. The most familiar (‘perfect’ or ‘complete’) form is for the two components to arise from adjacent insertions and be attached their whole length. However, in many instances the insertions of the two components are somewhat distant, so the organs are well separated at their base, but ω extends distally to meet acm and they conjoin for all or most of the length of the seta. The latter ‘imperfect’ (or ‘incomplete’) state, which Grandjean sometimes referred to as ‘maladjusted’, is probably a precursor and is found in the non-poronotic taxa listed above, as well as in what can be considered early-derivative Poronota, such as Phenopelopoidea (Unduloribatidae, Phenopelopidae) and Achipterioidea (Achipteriidae, Tegoribatidae).
Cymbaeremaeoidea lack the double-horn, while Licneremaeoidea exhibit all states of this character. There is an imperfect double-horn in Passalozetidae, Micreremidae, Dendroeremaeidae and some Adhaesozetidae (Phylleremus), and a perfect one in Adhaesozetes. In other licneremaeoid families there is no association of ω and acm. While Scutoverticidae are sometimes described as having an ‘incomplete double-horn’ (e.g. Schäffer et al. 2010, Pfingstl et al. 2010b), it seems unreasonable to use the term since the structures are not really conjoined for any part of their length: typically in this family, ω inclines distally and just touches acm with its tip.
Among Ameronothroidea, an association of acm and ω occurs only in Podacaridae, where all examined species have a perfect double-horn (Grandjean 1955, Ermilov et al. 2012). In Ameronothrus marinus ω is closely adjacent (not really attached) to non-eupathidial seta cm, behind it (Schubart 1975), but this unique arrangement reflects no homology with the double-horn. Podacaridae are the only entire family outside Poronota to have a perfect double-horn, but this has been overlooked in the past (Weigmann and Schulte 1977; Behan-Pelletier 1997, Norton and Behan-Pelletier 2009). It seems significant, especially in light of recent molecular studies that raise doubt about the monophyly of Ameronothroidea (Pfingstl 2017) and even infer that Podacaridae are nested within poronotic taxa (Schäffer et al. 2010). By contrast, there are groups in which the double-horn shows considerably homoplasy, as in Licneremaeoidea (see above) and Peloppiidae, where there may be no association (e.g. Parapyroppia), an imperfect double-horn (e.g. Metrioppia), or a perfect one (Ceratoppia).
20. Leg respiratory organs of adult – Respiratory porose organs occur on the legs of virtually all brachypyline oribatid mite adults, usually in the form of a surface porose area on all femora and on trochanters III and IV (‘trochantero-femoral system’), but they can occur also on tibiae and tarsi (‘tibio-tarsal system’). Various types of invaginated organs also are known, usually in taxa that are resistant to desiccation. Internalization of the respiratory surface as a saccule, brachytrachea (elongated, thick saccule) or trachea (fine tube, often with terminal bulb) may directly reduce water-loss, but also may allow increased waterproofing of the general cuticle (see reviews of Norton and Alberti 1997 and Norton et al. 1997 for explanations and citations).
In Cymbaeremaeoidea, internalization as saccules or brachytracheae is typical, though they can be mixed with porose areas according to species; in some, e.g. most Scapheremaeus species, only porose areas exist. Most Licneremaeoidea exhibit porose areas but internalized organs exist, according to taxon. Internalization of at least some organs is typical of Passalozetidae and Scutoverticidae, and some of the latter express the trochantero-femoral system as fine tracheae. Similarly, all forms of leg respiratory structures are known from Ameronothroidea. The fortuynioid families, Podacaridae and Chudalupia have only porose areas, and Ameronothrus species have only internalized organs (trochantero-femoral brachytracheae, and saccules on tibiae but not tarsi). The trochantero-femoral system of Aquanothrinae has either a mixed pattern of saccules and porose areas (P. spooneri) or only tracheae (all other species).
Without discussion, Weigmann and Schulte (1977) considered the presence of invaginated porose organs on legs to be derived within their broad concept of Ameronothridae and, taken in isolation, this is a reasonable conclusion from the seeming specialization of an internalized respiratory surface (e.g. Norton and Alberti 1997). In a slightly larger context, Behan-Pelletier (1988) considered invagination as saccules to be an ‘underlying synapomorphy of Cymbaeremaeidae + Ameronothridae + Scutoverticidae’, though with various expressions within the groups. But the distribution of these structures seems too great for this to be true. E.g., legs have saccules in some Ameroidea (Eremobelbidae) and tracheae in many Plateremaeoidea; more importantly, leg saccules and brachytracheae are found in most members of the earliest-derivative brachypyline families, Neoliodidae and Hermanniellidae. Therefore, we tentatively pose the new hypothesis that internalized surfaces evolved early in Brachypylina and therefore are plesiomorphic in adults of the three superfamilies. The expression of leg respiratory organs in adults as porose areas is considered a derived, paedomorphic transition, since that is their state in juveniles of these groups.
The early internalization of leg respiratory surfaces in Brachypylina may have been closely related to the development of the internalized apodemato-acetabular system (AAS), which characterizes Brachypylina (Norton et al. 1997). Both respiratory systems may have adapted to desiccating environments concurrently and in the same way. Regarding only the AAS, Norton and Alberti (1997) postulated that what started as a water-saving device may have had a great metabolic advantage, a ‘pre-adaptation’ (exaptation) that was key to the diversification of Brachypylina. But the invaginated leg respiratory surfaces—plesiomorphic in our scenario—may have become unimportant under the more humid environments in which most Brachypylina radiated, and reverted paedomorphically to simpler porose areas, whereas the AAS retained its metabolic importance in all environments.
21. Primilateral setae – The primilateral pair (pl) are fundamental, eustasic setae of tarsi, i.e. they are present beginning with the larva; if not formed then, they never appear during ontogeny. There is a diversity of patterns in macropyline groups, but in Brachypylina they occur only on tarsus Ia, and their absence is uncommon (Grandjean 1959). Their loss is clearly a regressive, apomorphic trait, but since they are eustasic it cannot be considered paedomorphic. Among Ameronothroidea, primilateral setae have been lost from Aquanothrinae and Ameronothrus, but retained in the fortuynioid families and Chudalupia. The pair is also retained in nearly all Podacaridae; the single exception known to us is Antarcticola meyeri, which has pl″ but lacks pl′. Among Cymbaeremaeoidea they have been lost from Cymbaeremaeus and nearly all examined Scapheremaeus (see Fernandez and Cleva 2010 for apparent exception) but retained in Ametroproctus. Among Licneremaeoidea they are lost from Micreremidae and Lamellareidae, lost (Adhaesozetes) or retained (Phylleremus) in Adhaesozetidae, and retained in Scutoverticidaeb, Passalozetidae, Charassobatidae and Dendroeremaeidae. Overall, the level of homoplasy is high, with both states present in each superfamily, and even within families (Cymbaeremaeidae, Adhaesozetidae) or genera (Scapheremaeus).
a Fernandez and Cleva (2010) indicated the presence of pl′ on tarsus III (their Fig. 6A) and pair (pl) on tarsus IV (their Fig. 6D) of a Scapheremaeus species, but we believe these are errors: the seta on tarsus III may be an antelateral seta, as the proximoventral setae labeled ‘(a)’ are probably (pv); the pair labeled ‘(pl)’ on tarsus IV are almost certainly (pv).
b In their analysis of leg setation in two species of Scutovertex, Ermilov et al. (2008; their Table 4 and Fig. 23) indicated that seta pl″ was absent from tarsus I, but this is an error (confirmed by S. Ermilov, personal communication, 2017): the pair exists, as in other members of the genus. They misinterpreted seta ft″ as a second solenidion in the larva, and pl″ was consequently misidentified as ft″. The solenidion added in the protonymph is therefore ω2, not ω3. Additionally, the label m is inadvertent in both the figure and table and should be deleted.
22. Primiventral setae of tarsi – The primiventral pair, (pv), are eustasic, fundamental setae that usually are present on all tarsi of brachypyline oribatid mites. Among Ameronothroidea, they show regression only in Aquanothrinae, where each genus has a different pattern: in Aquanothrus the only loss is on tarsus III, where pv″ is absent; in both Paraquanothrus species the pair is absent from tarsi II and III, pv′ is absent from I, and pv″ is absent from IV. In Cymbaeremaeoidea, regression is limited to Scapheremaeus, in which a variety of patterns of presence-absence has been reported among species. In Licneremaeoidea, primiventral setae show regression only in two families: in Micreremidae pv′ is absent from tarsi I and II, and pv″ is absent from tarsi III and IV; in Charassobatidae pv′ is absent from tarsus I, but the pair is present on II and perhaps on III (see Grandjean 1958a). While each of these taxa exhibits apomorphic regression in primiventral setae, the exact patterns differ.
Regression of primiventral setae can affect a commonly used character of juveniles: the setal-count formula for protonymphal leg IV. This leg has a highly regressed setation when it first forms in oribatid mites (Grandjean 1946a), but interpretations should go beyond numbers. In the typical formula of 0-0-0-0-7, the tarsal setae are ft″, (p), (u), and (pv), so if a primiventral seta is absent the formula is diminished. When represented only by counts, the 0-0-0-0-6 formula of Paraquanothrus, Micreremus and some Scapheremaeus (all lacking pv″) is indistinguishable from that of, e.g., Lohmanniidae, which have both primiventrals but lack one proral seta, p″, or that of some Epilohmanniidae, which lack pv″, but also lack pair (p) and develop the antelateral pair, (a), to result in the same formula.
23. Iteral setae – The iteral pair (it) of tarsal setae always are post-larval, appearing between the proral and tectal pairs. They also are amphistasic, i.e., their presence and instar of appearance vary among taxa, among the four tarsi, and rarely even between members of the pseudosymmetrical pair. The diverse ontogenetic patterns were summarized by Grandjean (1961, 1964a), whose conceptual model involved evolutionary trends in which ‘resistance’ to regression (i.e., to ontogenetic delay and eventual failure to form) of iteral setae decreases from leg I to IV. Among Brachypylina iteral setae are added in the tritonymph (n3) or adult (Ad) or fail to form (0), and both the most plesiomorphic (n3-n3-n3-n3) and most apomorphic (0-0-0-0) formulas can be found in early-derivative groups (some Neoliodoidea and all Hermannielloidea, respectively). Ameronothroidea have a diversity of formulas, including: the rare n3-n3-n3-Ad (Ameronothrus) which is the most plesiomorphic for the groups in question; the slightly more regressive and very common n3-n3-n3-0 (Podacaridae, Tegeocranellidae, Fortuyniidae, Chudalupia a); the rare n3-n3-0-0 (Selenoribatidae); and the highly apomorphic 0-0-0-0 (Aquanothrinae). Cymbaeremaeoidea have the common n3-n3-n3-0 (Ametroproctus, Cymbaeremaeus some Scapheremaeus speciesb) or 0-0-0-0 (some Scapheremaeus species). Most Licneremaeoidea have the common n3-n3-n3-0 (Scutoverticidae, Dendroeremaeidae Micreremidaec), but Adhaesozetidae have the derived formulas n3-n3-0-0 (Phylleremus) and 0-0-0-0 (Adhaesozetes). Charassobatidae have the unique formula Ad-Ad-[0, Ad]-0. Ontogeny of iterals is unknown in Lamellareidae and Passalozetidae, but they probably follow the common formula, being present on adult tarsi I-III and absent from IVd.
Weigmann and Schulte (1977) restricted their analysis of iteral setae to adult tarsus IV. By apparently following Grandjean’s concepts, they considered the presence of iterals in Ameronothrus to be plesiomorphic and their absence in other analysed taxa to be apomorphic; the latter was considered a weak trait, implying that losses might be convergent. After expanding the iteral seta character to an ontogenetic formula (for all legs and for an expanded list of taxa), we agree with their view. The complete loss of iteral setae is unusual in Brachypylina: Grandjean (1961, 1964a) knew it only in Hermannielloidea and some Zetorchestidae. Aquanothrinae are the only Ameronothroidea to have this trait, but complete regression also has occurred in Cymbaeremaeoidea (some Scapheremaeus) and Licneremaeoidea (Adhaesozetes) as well, so homoplasy weakens it as an informative character state. As it seems to result from ontogenetic delays operating in multiple clades, it is a paedomorphic trait, prone to convergence.
a Wallwork (1981) wrote that Chudalupia meridionalis was similar to Ameronothrus in having iteral setae on all tarsi of adults, but this appears to be an error. We studied multiple adult and tritonymph topotypes of C. meridionalis and all possessed iterals on tarsi I-III while lacking them on tarsus IV.
b For Micreremus, we follow Grandjean (1961), who assigned the common formula n3-n3-n3-0. By contrast, Pfingstl and Krisper (2011b) indicated in their Table 1 that the iteral pair forms on all legs of M. brevipes (Michael, 1888), and on leg IV they appear uniquely early, in the deutonymph when the tectal pair usually appear. These are inadvertent errors (T. Pfingstl, personal communication, 2017): their Fig. 15 shows the correct setal labeling for the deutonymph: possessing tectals and lacking iterals (which never form on tarsus IV). Their Table 1 shows the correct addition of the iteral pair to the tritonymph on tarsus III, but their illustration of the adult leg III (Fig. 20) shows a different interpretation, lacking iteral setae but including a pair of fastigial setae that are absent from Table 1. We have not studied the ontogeny of this species but based on a study of the adults we believe the table is correct. An unrelated error in their Table 1 is the inclusion of seta d on genu I: instead it should be l′, as they correctly list for genu II – genual seta d forms only on leg IV of this species.
c According to Fernandez and Cleva (2009), one species of Scapheremaeus purportedly has (it) on all tarsi, but we question the accuracy and labeling of certain illustrations, particularly their Fig. 8. Most importantly, the captions of Fig. 8B (leg II) and 8D (leg IV) may have been inverted. There is a solenidion on the tarsus in Fig. 8D, but tarsus IV never has a solenidion in oribatid mites. Similarly, the tarsus in Fig. 8B lacks a solenidion, but one is always present on tarsus II. Also, the shapes of the respective femora are consistent with a numbering error. Questionable notations include the labeling of a setal pair on Fig. 8B as primilaterals, which only exist on tarsus I (see Remark 21 above), the labeling of a dorso-distal tarsal pair on Fig. 8D as (v), and the supposed absence of unguinal setae.
d Martínez and Herrero (2006) used the label ‘(it)’ on tarsi I and IV of an Argentinian species of Passalozetes, but their illustration is inconclusive and we doubt the accuracy of the labeling; for example, neither fastigials nor tectals are identified on leg I, and no proral setae were identified on leg IV, all of which are almost universally present in Brachypylina.
24. Proximal accessory setae of tarsus I – Post-larval instars of oribatid mites commonly add setae in lateral and ventral rows toward the proximal end of the segment, and unlike the iteral pair these are eustasic setae (Grandjean 1958b, therein called ‘posterior’ accessory setae; see also Norton and Sidorchuk 2014). While macropyline groups can have numerous setae in these rows, they are notably regressed in Brachypylina. It is common in Brachypylina for tarsus I to add l″ and v′ in the adult but they often are absent in the taxa relevant here (Behan-Pelletier 1988), as well as many other superfamilies. No known member of Ameronothroidea adds accessory setae. Among Cymbaeremaeoidea both l″ and v′ are added in Cymbaeremaeus and Ametroproctus, but neither appears in Scapheremaeus a. In Licneremaeoidea one or two proximal accessory setae are added in Passalozetidae and Dendroeremaeidaeb, but not in Scutoverticidae, Charassobatidae, Micreremidae or Lamellareidae; Adhaesozetidae is mixed (present in Adhaesozetes, absent from Phylleremus). Overall, the apomorphic, regressive loss of these setae seems common and homoplasious.
a Fernandez and Cleva (1997; their Fig. 4A) illustrated an unlabeled seta proximal to pair (pv) on tarsus IV of a Scapheremaeus species. The ontogeny is unknown, but the seta seems unlikely to be a proximal accessory seta, as there is no example of such setae on tarsus IV of any Brachypylina. Also, since no other tarsus of this species has an accessory seta, the presence of one on tarsus IV would represent a priority opposite from that proposed by Grandjean (1958b): in phylogenetic time, posterior accessory setae regress first on tarsus IV, and last on tarsus I. The seta may have been an anomaly.
b Behan-Pelletier et al. (2005, their Table 1) reported a unique pattern of proximal accessory setae in Dendroeremaeus krantzi Behan-Pelletier, Eamer and Clayton, 2005: three setae are indicated as forming in the adult—(v) on tarsus I and v′ on tarsus II. After studying their illustrations (which are correct), and paratype specimens, we determined instead that seta l″ forms in the adult on tarsus I, and no seta is added on tarsus II; i.e., there is nothing unusual about the ontogeny.
25. Subunguinal seta of tarsus I – Unpaired seta s is eupathidial in most oribatid mite adults, and this probably is the ancestral state in Brachypylina, being true of most macropyline groups as well as the early-derivative superfamily Hermannielloidea (but not Neoliodoidea). In the three superfamilies of concern here, s is usually a normal, solid seta; this can be interpreted as a derived, paedomorphic trait, since s becomes eupathidial during ontogeny in most oribatid mites. Behan-Pelletier (1988) considered the normal form of s to be a synapomorphy of Cymbaeremaeoidea, Ameronothroidea and Licneremaeoidea (except Charassobatidae). However, there is considerable homoplasy, as s is eupathidial in some Licneremaeoidea (e.g., Scutovertex, Dendroeremaeidae, Charassobatidae) and Ameronothroidea (Tegeocranellidae).
26. Retention of coupled-seta d on genua I-III and tibiae II-IV of adult – In Brachypylina, the close coupling of seta d with the respective solenidion on genua I-III and on all tibiae is a plesiomorphic trait, typical also of Mixonomata and Nothrina. Grandjean (1964b) noted that genu IV lacks a solenidion, with exceptions in Neoliodes (Neoliodoidea)a, and d is consistently present. Successive derivations involve the regression in size or complete loss of seta d from the adult (only), various partial losses, and the loss of d from all instars (Grandjean 1946a, 1954a). In most cases, genua I-III and tibiae II-IV exhibit similar states, but that of tibia I is more regressive (d is rarely present in the adult, even if other segments retain it) and is not considered here.
Seta d is present in juveniles but lost in the adult of most Cymbaeremaeoidea, although the pattern is abnormal in some Ametroproctus species, being lost from some segments but retained on others (Behan-Pelletier 1988). Among Licneremaeoidea, seta d is present in juveniles but lost in adults in some families (Scutoverticidae, Licneremaeidae), but absent from all instars in others (Micreremidae, Adhaesozetidae, Dendroeremaeidae). Among Ameronothroidea, d is absent from all instars in the fortuynioid families, but d is present in juveniles of the other families. Seta d is lost from adults of Podacaridae and Chudalupia. In Ameronothrus the genua retain d in the adult; tibiae may retain or lose it, according to species. Among Aquanothrinae, Paraquanothrus species lose d in the adult, but the pattern is mixed in Aquanothrus. In A. montanus, d is retained on genua I and II but lost from genu III; d is lost from tibiae I-III but retained on tibia IV (the Botswana species also loses d from tibia IV). The unique Aquanothrus pattern represents an intermediate level of regression in seta d that has been overlooked or misinterpreted, due to original observational errors (see above).
We agree with Weigmann and Schulte (1977) who considered the retention of d to be plesiomorphic in Ameronothroidea. Behan-Pelletier (1988) considered it apomorphic in some species of the cymbaeremaeoid genus Ametroproctus; she did not explain this, but may have envisioned it as a reversal, since taxa that she considered outgroups lose d in the adult. Regardless, the confusing distribution of states shows this character to be highly homoplasious.
a Supposed examples in two more-derived families—Cymbaeremaeidae and Lamellareidae—were reported by later authors, but each is incorrect. The single dorsal setiform organ on genu IV of Scapheremaeus species has been considered (and illustrated as) a solenidion by multiple authors, including us (Travé and Fernandez 1986; Fernandez and Cleva 2009; Norton et al. 2010; Ermilov and Minor 2015b). We reexamined this structure in Scapheremaeus rodickae Norton, Franklin and Crossley, 2010, as well as seven other Scapheremaeus species in our collections and in each case there is no solenidion, but rather a short, smooth seta d, with much the same form as the solenidion on genua I-III. It is distinguishable from a solenidion by: the presence of pigment, the apobasic alveolus, the lack of a central canal and the presence of birefringence in polarized light. Based on a large ongoing study of Scapheremaeus, Dr. N. Fernandez confirmed (personal communication, 2017) that genu IV never has a solenidion. Similarly, Martínez et al. (1995) reported and illustrated a dorsal solenidion on genu IV of Tenuelamellarea argentinensis. Dr. P. Martínez kindly reexamined his mites and confirmed the error (personal communication, 2017): the structure is instead seta d.
27. Solenidion ω2 of tarsus II – Most Brachypylina develop a second solenidion (ω2) on tarsus II during ontogeny, usually in the deutonymph. To Grandjean (1964b) the loss of ω2 characterized a second ‘accrochage’, a sticking- or holding-point in the regressive evolution of solenidia. Regression appears to have reached this point in about a dozen genera of diverse families, and further losses are rare in Brachypylina. Among Ameronothroidea ω2 is absent from Tegeocranellidae and Selenoribatidae, but retained by Ameronothrus, Chudalupia and Podacaridae. Most Fortuyniidae have lost ω2, but some Fortuynia retain it. Aquanothrinae also are mixed, with ω2 being lost from Paraquanothrus but retained by Aquanothrus. Among Cymbaeremaeoidea, ω2 is absent from Scapheremaeus and Cymbaeremaeus a, but has been retained in Ametroproctus. Among Licneremaeoidea, ω2 is absent from Micreremidae, Lamellareidae, Charassobatidae and some Adhaesozetidae (Adhaesozetes); it is retained in other Adhaesozetidae (Phylleremus) and in Scutoverticidae and Dendroeremaeidae. The absence of ω2 is a regressive, paedomorphic loss that appears to be homoplasious, having occurred independently multiple times, even within families and genera.
a Pfingstl and Krisper (2011a; their Table 2) indicated that solenidion ω2 is added to tarsus II of Cymbaeremaeus cymba in the deutonymph, but this seems to be an error. In another paper of the same year (Pfingstl and Krisper 2011b; their Table 3) they indicated its absence, which is consistent with Grandjean (1964b) and our own studies of C. cymba specimens from Spain.
28. Coupling of fastigial setae with solenidion ω1 (= ω) on tarsus II – On tarsus I of Brachypylina it is common for fastigial seta ft″ to be positioned close to a solenidion—most often this is ω1, or both ω1 and ω2—and they sometimes share an alveolus. Often the famulus joins these, in what has been called the ‘tarsal cluster’ (Grandjean 1935). Coupling of fastigial setae and solenidia is less common on tarsus II. On tarsus II of Paraquanothrus (which lack ω2) ft′ is coupled with ω (i.e., ω1) in the same alveolus. In Aquanothrus both fastigial setae are isolated, while ω1 is adjacent instead to ω2. Ameronothrus shows the latter state in some species; in others ft′ is closely coupled with ω1 and ft″ with ω2. Among Podacaridae, Podacarus exhibits a similar ft′-ω1 and ft″-ω2 coupling, as does Pseudantarcticola georgiae (Wallwork, 1970); by contrast, in Antarcticola meyeri it is ft″ that couples with ω1. In Halozetes, ft′ is coupled with ω1 or independent, according to speciesa. Both fastigials are independent in the fortuynioid families. We know of no coupling in either Cymbaeremaeoidea or Licneremaeoidea, but such details are rarely reported or interpretable from figures.
a Coetzee (2000; her Fig. 7) illustrated ft′ as coupled to ω2 on tarsus II of Halozetes belgicae mickii Coetzee, 2000. The ontogeny of this subspecies was not described, but we believe the coupled solenidion is actually ω1; it is the more proximal of the two, and in all Podacaridae for which ontogeny is known, ω1 is proximal to ω2.
This study was made possible through the kind assistance of the following people. Drs. Tim Graham (U.S. Geological Survey, Moab, Utah) and John D. Spooner (Department of Biology & Geology, University of South Carolina, Aiken) collected and donated specimens of Paraquanothrus grahami and P. spooneri, respectively, and provided essential information. Specimens of Aquanothrus montanus from South Africa were donated by Drs. C. M. Engelbrecht and Louise Coetzee, (the National Museum, Bloemfontein, Republic of South Africa) and they (along with Dr. Lizel Hugo-Coetzee) also provided helpful information. Aquanothrus specimens from Botswana were donated by Dr. Merlijn Jocqué (Laboratory of Aquatic Ecology, Leuven, Belgium). Topotype specimens of Chudalupia meridionalis were donated by Dr. Ian A. E. Bayly (Monash University, Australia). Scanning and transmission micrographs of P. grahami (some previously published in Norton et al. 1996b) were provided by the late Prof. Dr. Gerd Alberti (University of Greifswald, Germany) and John Gardner (Brigham Young University, Utah). Drs. Sergey Ermilov (Tyumen State University, Russia), Nestor Fernandez (Universidad Nacional de La Pampa, Argentina) and Pablo Martínez (Universidad Nacional de Mar del Plata, Argentina) kindly responded to questions about their publications. Dr. Diego Fontaneto (Istituto per lo Studio degli Ecosistemi, Verbania Pallanza, Italy) confirmed the generic identity of the rotifer Philodina sp. Prof. D. A. Crossley (University of Georgia, Athens) provided working facilities for author E. Franklin. Dr. Tobias Pfingstl (University of Graz, Austria) allowed us to see his 2017 paper before publication, kindly responded to several inquiries, and reviewed the manuscript. Dr. Valerie Behan-Pelletier (Agriculture and Agri-Food Canada, Ottawa) provided critiques of both early and late manuscript drafts, as well as specimens and information about some relevant species. The thoughtful and detailed attention of Drs. Behan-Pelletier, Pfingstl and editor Dr. Ekaterina Sidorchuk were particularly helpful. We are grateful to all for their generosity and interest.
Alberti G. 1979. Fine structure and probable function of genital papillae and Claparéde organs of Actinotricha. In: Rodriguez J.G. (Ed.). Recent Advances in Acarology, vol. 2. New York: Academic Press, Inc. p. 501-507. doi:10.1016/B978-0-12-592202-9.50072-0 ![]()
Alberti G., Coons B. 1999. Acari – Mites. In: Harrison F.W., Foelix R.F. (Eds). Microscopic anatomy of invertebrates. Volume 8C: Chelicerate Arthropoda. New York: Wiley-Liss. p. 515-1265.
Alberti G., Fernandez N.A. 1990. Aspects concerning the structure and function of the lenticulus and clear spot of certain oribatids (Acari, Oribatida). Acarologia, 31: 65-72.
Alberti G., Moreno-Twose A.I. 2012. Fine structure of the primary eyes in Heterochthonius gibbus (Oribatida, Heterochthoniidae) with some general remarks on photosensitive structures in oribatid and other actinotrichid mites. Soil Organisms, 84: 391-408.
Alberti G., Moreno A.I., Kratzmann M. 1994. The fine structure of trichobothria in moss mites with special emphasis on Acrogalumna longipluma (Berlese, 1904) (Oribatida, Acari, Arachnida). Acta Zoologica, Stockholm, 75: 57-74. doi:10.1111/j.1463-6395.1994.tb00962.x ![]()
Balogh J. 1961. Identification keys of world oribatid (Acari) families and genera.- Acta Zoologica Academiae Scientiarum Hungaricae, 7: 243-344.
Balogh J. 1963. Identification keys of holarctic oribatid mites (Acari) families and genera. Acta Zoologica Academiae Scientiarum Hungaricae, 9: 1-60.
Balogh J. 1972. The oribatid genera of the world. Budapest: Akademiai Kiadó. pp. 188.
Balogh J., Balogh P. 1992. The oribatid mites genera of the world. vol. 1. Budapest: The Hungarian National Museum Press. pp. 263.
Balogh J., Mahunka S. 1981. New data to the knowledge of the oribatid fauna of Neogaea (Acari) VI.- Acta Zoologica Academiae Scientiarum Hungaricae, 27: 49-102.
Baranek S.E. 1981. Contribución al estudio de la familia Oripodidae (Acari, Oribatei). I. Physis, Secc. C, Buenos Aires, 40(98): 19-25.
Bayly I.A.E. 1997. Invertebrates of temporary waters in gnammas on granite outcrops in Western Australia. Journal of the Royal Society of Western Australia, 80: 167-172.
Behan-Pelletier V.M. 1987. Redefinition of Ametroproctus (Acari: Oribatida) with descriptions of new species. Canadian Entomologist, 119: 505-535. doi:10.4039/Ent119505-6 ![]()
Behan-Pelletier V.M. 1988. Systematic relationships of Ametroproctus with modified definition of Cymbaeremaeidae (Acari: Oribatida). In: Channabasavanna G.P., Viraktamath C.A. (Eds). Progress in Acarology. vol. 1. New Delhi: Oxford & IBH Publishing Co. Pvt. Ltd. p. 301-307.
Behan-Pelletier V.M. 1989. Limnozetes (Acari: Oribatida: Limnozetidae) of Northeastern North America. Canadian Entomologist, 121: 453-506. doi:10.4039/Ent121453-6 ![]()
Behan-Pelletier V.M. 1991. Observations on genital papillae of pycnonotic Brachypylina (Acari: Oribatida). Acarologia, 32: 71-78.
Behan-Pelletier V.M. 1997. The semiaquatic genus Tegeocranellus (Acari: Oribatida: Ameronothroidea) of North and Central America. Canadian Entomologist, 129: 537-577. doi:10.4039/Ent129537-3 ![]()
Behan-Pelletier V., Eamer B. 2007. Aquatic Oribatida: adaptations, constraints, distribution and ecology. In: Morales-Malacara J.B., Behan-Pelletier V., Ueckermann E., Pérez T.M., Estrada-Venegas E.G., Badil M. (Eds). Acarology XI: Proceedings of the International Congress. México: Universidad Nacional Autónoma de México. p. 71-82.
Behan-Pelletier V.M., Walter D.E. 2007. Phylleremus n.gen. from leaves of deciduous trees in eastern Australia (Oribatida: Licneremaeoidea). Zootaxa, 1386: 1-17.
Behan-Pelletier V.M., Eamer B., Clayton M. 2005. Dendroeremaeidae n. fam., from forest trees in Western North America (Acari: Oribatida: Licneremaeidae). Acarologia, 45: 321-339.
Bernini F. 1984. Notulae Oribatologicae XXXIII. New records of Cryptacarus promecus Grandjean and Cryptogalumna cryptodonta for Italy (Acarida, Oribatida). Animalia, Catania, 11: 143-153.
Chan M.A., Moser K., Davis J.M., Southam G., Hughes K., Graham T. 2005. Desert potholes: Ephemeral aquatic microsystems. Aquatic Geochemistry, 11: 279-302. doi:10.1007/s10498-004-6274-8 ![]()
Coetzee L. 1987. The South African Lamellareidae Balogh, 1972 (Acari, Oribatei). Navorsinge van die Nasionale Museum, Bloemfontein, 5: 326-353.
Coetzee L. 2000. A new subspecies of Halozetes belgicae (Acari, Oribatida, Ameronothridae) from the marine supra-littoral of the sub-Antarctic Marion Island. Acarologia, 40: 439-447.
Colloff M.J. 2009. Comparative morphology and species-groups of the oribatid mite genus Scapheremaeus (Acari: Oribatida: Cymbaeremaeidae), with new species from South Australia. Zootaxa, 2213: 1-46.
Colloff M.J. 2010. The hyperdiverse oribatid mite genus Scapheremaeus (Acari: Oribatida: Cymbaeremaeidae) in Australia, with descriptions of new species and consideration of biogeographical affinities. Zootaxa, 2475: 1-38.
Colloff M.J., Halliday R.B. 1998. Oribatid mites: a catalogue of the Australian genera and species. Monographs on Invertebrate Taxonomy, vol. 6. Melbourne: CSIRO Publishing. pp. 224.
Dabert M., Witalinski W., Kazmierski A., Olszanowski Z., Dabert J. 2010. Molecular phylogeny of acariform mites (Acari, Arachnida): Strong conflict between phylogenetic signal and long-branch attraction artifacts. Molecular Phylogenetics and Evolution, 56: 222-241. doi:10.1016/j.ympev.2009.12.020 ![]()
Engelbrecht C.M. 1974. The genus Halozetes (Oribatei: Acari) on Marion Island. Navorsinge van die Nasionale Museum, Bloemfontein, 3: 1-25.
Engelbrecht C.M. 1975. New ameronothroid (Oribatei: Acari) taxa from the Republic of South Africa and the Islands Gough and Marion. Navorsinge van die Nasionale Museum, Bloemfontein, 3: 53-88.
Ermilov S.G., Minor M.A. 2015a. New species of oribatid mites (Acari: Oribatida) of the genera Austrachipteria (Achipteriidae), Cultroribula (Astegistidae) and Microlamellarea (Lamellareidae) from New Zealand. Biologia, 70: 1501-1519. doi:10.1515/biolog-2015-0173 ![]()
Ermilov S.G., Minor M.A. 2015b. The genus Scapheremaeus (Acari, Oribatida, Cymbaeremaeidae) in the oribatid mite fauna of New Zealand, with description of two new species. ZooKeys, 508: 69-83. doi:10.3897/zookeys.508.10005 ![]()
Ermilov S.G., Łochyńska M., Olszanowski Z. 2008. The cultivation and morphology of juvenile stages of two species from genus Scutovertex (Acari: Oribatida: Scutoverticidae). Annales Zoologici (Warszawa), 58: 433-443. doi:10.3161/000345408X326762 ![]()
Ermilov S.G., Stary J., Block W. 2012. Morphology of juvenile instars of Ameronothridae (Acari: Oribatida). Zootaxa, 3224: 1-40.
Ermilov S.G., Tolstikov A.V., Salavatulin V.M., Bragin E.A. 2015. Morphology of juvenile stages in two species of arboreal oribatid mites, Scapheremaeus palustris and Phauloppia nemoralis (Acari, Oribatida). Zoologicheskii Zhurnal, 94: 26-36. (in Russian; English version: Entomological Review, 95: 126-136). doi:10.1134/S0013873815010145 ![]()
Fernandez N.A., Cleva R. 1997. Contribution à la connaissance du genre Scapheremaeus habitant les plantes epiphytes. I. Scapheremaeus tillandsiae sp.n. Acarologia, 38: 289-296.
Fernandez N.A, Cleva R. 2009. Contribution à la connaissance des oribates d'Argentine. I. Le genre Scapheremaeus: Scapheremaeus chaquensis n. sp. Acarologia, 49: 55-67.
Fernandez N., Cleva R. 2010. Une nouvelle espèce de Scapheremaeus (Arachnida, Acari, Oribatida, Cymbaeremaeidae) de Madagascar: Scapheremaeus pauliani n. sp. Zoosystema, 32: 101-115. doi:10.5252/z2010n1a5 ![]()
Fernandez N.A., Marcangeli J., Eguaras M. 1997. Les Acariens (Oribates) des zones arides d'Argentine. II. Huilicheremaeus michaii, n. gen., n. sp. Acarologia, 38: 79-85.
Fuangarworn M., Norton R.A. 2013. Psammochthoniidae n. fam., a paedomorphic family of oribatid mites (Oribatida: Enarthronota) from sandy soil in Thailand, Brazil and the USA. Zootaxa, 3691: 473-499. doi:10.11646/zootaxa.3691.4.7 ![]()
Fujikawa T. 1991. List of oribatid families and genera of the world. Edaphologia, 46: (i-vii) 1-132.
Graham T.B., Norton R.A. 1998. Uncharismatic microfauna of the Colorado Plateau: Notes on distribution and ecology of an undescribed, pothole-dwelling ameronothroid (Acari: Ameronothridae) mite. In: Hill L.M. (Ed.). Learning from the Land: Grand Staircase-Escalante National Monument Science Symposium Proceedings. Salt Lake City: U.S. Department of the Interior, Bureau of Land Management. p. 477-483.
Grandjean F. 1934a. La notation des poils gastronotiques et des poils dorsaux du propodosoma chez les Oribates (Acariens). Bulletin de la Société zoologique de France, 59: 12-44.
Grandjean F. 1934b. Les poils des épimères chez les Oribates (Acariens). Bulletin du Muséum National d'Histoire Naturelle, 2e série, 6: 504-512.
Grandjean F. 1935. Les poils et les organes sensitifs portés par les pattes et le palpe chez les Oribates. Bulletin de la Société zoologique de France, 60, 6-39.
Grandjean F. 1946a. La signification evolutive de quelques caractères des Acariens (1re série). Bulletin Biologique de la France et de la Belgique, 79 : 297-325.
Grandjean F. 1946b. Les poils et les organes sensitifs portés par les pattes et le palpe chez les Oribates. Troisième partie. Bulletin de la Société zoologique de France, 71: 10-29.
Grandjean F. 1947a. Observations sur les Oribates (17e série). Bulletin du Muséum National d'Histoire Naturelle, 2e série, 19: 165-172.
Grandjean F. 1947b. Observations sur les Oribates (18e série). Bulletin du Muséum National d'Histoire Naturelle, 2e série, 19: 395-402.
Grandjean F. 1948. Sur les Hydrozetes (Acariens) de l'Europe occidentale. Bulletin du Muséum National d'Histoire Naturelle, 2e série, 20: 328-335.
Grandjean F. 1949a. Formules anales, gastronotiques, génitales et aggénitales du développement numériques des poils chez les Oribates. Bulletin de la Société Zoologique de France, 74: 201-225.
Grandjean F. 1949b. Observation et conservation des très petits arthropodes. Bulletin du Muséum National d'Histoire Naturelle, 2e série, 21: 363-370.
Grandjean F. 1951. Comparaison du genre Limnozetes au genre Hydrozetes (Oribates). Bulletin du Muséum National d'Histoire Naturelle, 2e série, 23: 200-207.
Grandjean F. 1952. Au sujet de l'ectosquelette du podosoma chez les Oribates superieurs et de sa terminologie. Bulletin de la Société zoologique de France, 77: 13-36.
Grandjean F. 1954a. Essai de classification des Oribates (Acariens). Bulletin de la Société Zoologique de France, 78: 421-446.
Grandjean F. 1954b. Etude sur les Palaeacaroides (Acariens, Oribates). Mémoires du Muséum National d'Histoire Naturelle (n.s), série A, Zoologie, 7: 179-274.
Grandjean F. 1955. Sur un Acarien des iles Kerguelen. Podacarus Auberti (Oribate). Mémoires du Muséum National d'Histoire Naturelle (n.s), série A, Zoologie, 8: 109-150.
Grandjean F. 1956a. Galumnidae sans carènes lamellaires (Acariens, Oribates), 1re série. Bulletin de la Société zoologique de France, 81: 134-150.
Grandjean F. 1956b. Sur deux espéces nouvelles d'Oribates (Acariens) apparentées à Oripoda elongata Banks 1904. Archives de Zoologie Expérimentale et Générale, 93: 185-218.
Grandjean F. 1957. L'infracapitulum et la manducation chez les Oribates et d'autres Acariens. Annales des Sciences Naturelles. Série 2. Zoologie, 19: 234-281.
Grandjean F. 1958a. Charassobates cavernosus Grandjean 1929 (Acarien, Oribate). Mémoires du Muséum National d'Histoire Naturelle (n.s), série A, Zoologie, 16: 121-140.
Grandjean F. 1958b. Sur le comportement et la notation des poils accessoires posterieurs aux tarses des Nothroides et d'autres Acariens. Archives de Zoologie Expérimentale et Générale, 96: 277-308.
Grandjean F. 1959. Observations sur les Oribates (40e série). Bulletin du Muséum National d'Histoire Naturelle, 31: 359-366.
Grandjean F. 1960. Damaeus arvernensis n. sp. (Oribate). Acarologia, 2: 250-275.
Grandjean F. 1961. Nouvelles observations sur les Oribates (1re série). Acarologia, 3: 206-231.
Grandjean F. 1962. Le genre Tegeocranellus Berl. 1913 (Oribates). Acarologia, 4: 78-100.
Grandjean F. 1963a. La néotrichie du genre Tricheremaeus d'après T. nemossensis n. sp. (Oribate). Acarologia, 5: 407-437.
Grandjean F. 1963b. Concernant Sphaerobates gratus, les Mochlozetidae et les Ceratozetidae (Oribates). Acarologia, 5: 284-305.
Grandjean F. 1963c. Sur deux espèces de Brachychthoniidae et leur développement (Oribates). Acarologia, 5: 122-151.
Grandjean F. 1964a. Nouvelles observations sur les Oribates (3e série). Acarologia, 6: 170-198.
Grandjean F. 1964b. La solénidiotaxie des Oribates. Acarologia, 6: 529-556.
Grandjean F. 1965a. Oribates mexicains (2e série). Stelechobates megalotrichus n.g., n.sp. Acarologia, 7: 532-563.
Grandjean F. 1965b. Fosseremus quadripertitus nom.nov. (Oribate). Acarologia, 7: 343-375.
Grandjean F. 1966. Selenoribates mediterraneus n.sp. et les Selenoribatidae (Oribates). Acarologia, 8: 129-154.
Grandjean F. 1968. Schusteria littorea n.g., n.sp. et les Selenoribatidae (Oribates).Acarologia, 10: 116-150.
Hammen L. van der 1963. Description of Fortuynia yunkeri nov. spec. and notes on the Fortuyniidae nov. fam. (Acarida, Oribatei). Acarologia, 5: 152-167.
Hammen L. van der 1980. Glossary of acarological terminology. Volume I. General terminology. The Hague: Dr. W. Junk Publ. pp. 244.
Hammer M. 1966. Investigations on the oribatid fauna of New Zealand. Part I. Biologiske Skrifter Det Kongelige Danske Videnskabernes Selskab, 15: 1-108.
Hammer M. 1971. On some oribatids from Viti Levu, Fiji Islands. Biologiske Skrifter Det Kongelige Danske Videnskabernes Selskab, 16: 1-60.
Iordansky S.N., Stein-Margolina V.A. 1993. Fine structure of the cuticle in nymphs and adults of oribatid mites (Acari, Cryptostigmata) with wrinkled preimaginal stages. Acarina, 1: 27-50.
Iseki A., Karasawa S. 2014. First record of Maculobates (Acari: Oribatida: Liebstadiidae) from Japan, with a redescription based on specimens from the Ryukyu Archipelago. Species Diversity, 19: 59-69. doi:10.12782/sd.19.1.059 ![]()
Jocqué M., Martens K., Riddoch B., Brendonck L. 2006. Faunistics of ephemeral rock pools in southeastern Botswana. Archiv für Hydrobiologie, 165: 415-431 doi:10.1127/0003-9136/2006/0165-0415 ![]()
Jocqué M., Riddoch B.J., Brendonck L. 2007. Successional phases and species replacements in freshwater rock pools: towards a biological definition of ephemeral systems. Freshwater Biology, 52: 1734-1744. doi:10.1111/j.1365-2427.2007.01802.x ![]()
Jocqué M., Vanschoenwinkel B., Brendonck L. 2010. Freshwater rock pools: a review of habitat characteristics, faunal diversity and conservation value. Freshwater Biology, 55: 1587-1602. doi:10.1111/j.1365-2427.2010.02402.x ![]()
Kok O.B. 1968. Lamellarea, a new oribatid genus (Acari) from South Africa. Acarologia, 10: 711-724.
Krause A., Pachl P., Schulz G., Lehmitz R. Seniczak A., Schaefer I., Scheu S. Maraun M. 2016. Convergent evolution of aquatic life by sexual and parthenogenetic oribatid mites. Experimental and Applied Acarology, 70: 439-453. doi:10.1007/s10493-016-0089-3 ![]()
Luxton M. 1985. Cryptostigmata (Arachnida: Acari) – a concise review. Fauna of New Zealand, 7: 1-106.
Maraun M., Erdmann G., Schulz G., Norton R.A., Scheu S., Domes K. 2009. Multiple convergent evolution of arboreal life in oribatid mites indicates the primacy of ecology.Proceedings of the Royal Society B, 276: 3219-3227. doi:10.1098/rspb.2009.0425 ![]()
Martínez P.A., Herrero A.C. 2006. A new species of Passalozetes (Acari: Oribatida) from the Andes mountains of Argentina. Zootaxa, 1160: 49-55.
Martínez P.A., Velis G.J., Eguaras M.J., Fernandez N.A. 1995. La famille Lamellareidae dans la République Argentine. Tenuelamellarea argentinensis n.sp. Acarologia, 36: 355-361.
McCullough E., Krisper G. 2013. Morphological analysis of the oribatid mite species Scutovertex pannonicus Schuster and description of its juvenile stages (Acari: Oribatida: Scutoverticidae). Zootaxa, 3619: 201-245. doi:10.11646/zootaxa.3619.3.1 ![]()
Messner B., Adis J. 1992. Die Plastronatmung bei aquatischen und flutresistenten terrestrischen Arthropoden (Acari, Diplopoda und Insecta). Mitteilungen der Deutschen Gesellschaft für allgemeine und angewandte Entomologie, 8: 325-327.
Minor M.A. 2011. Spatial patterns and local diversity in soil oribatid mites (Acari: Oribatida) in three pine plantation forests. European Journal of Soil Biology, 47: 122-128. doi:10.1016/j.ejsobi.2011.01.003 ![]()
Mortimer E., Jansen van Vuuren B., Lee J.E., Marshall D.J., Convey P., Chown S.L. 2011. Mite dispersal among the Southern Ocean Islands and Antarctica before the last glacial maximum. Proceedings of the Royal Society of London, B, Biological Sciences, 278: 1247-1255. doi:10.1098/rspb.2010.1779 ![]()
Norton R.A. 1998. Morphological evidence for the evolutionary origin of Astigmata (Acari: Acariformes). Experimental and Applied Acarology, 22: 559-594. doi:10.1023/A:1006135509248 ![]()
Norton R.A., Alberti G. 1997. Porose integumental organs of oribatid mites (Acari, Oribatida). 3. Evolutionary and ecological aspects. Zoologica, Stuttgart, 146: 115-143.
Norton R.A., Behan-Pelletier V.M. 1986. Systematic relationships of Propelops, with a modification of family-group taxa in Phenopelopoidea (Acari: Oribatida). Canadian Journal of Zoology, 64: 2370-2383. doi:10.1139/z86-353 ![]()
Norton R.A., Behan-Pelletier V.M. 2009. Chapter 15, Oribatida. In: Krantz G. W., Walter D. E. (Eds). A Manual of Acarology. Lubbock, Texas: Texas Tech University Press. p. 430-564.
Norton R.A., Kethley J.B. 1994. Ecdysial cleavage lines of acariform mites (Arachnida, Acari). Zoologica Scripta, 23: 175-191. doi:10.1111/j.1463-6409.1994.tb00383.x ![]()
Norton R.A., Lions J.C. 1992. North American Synichotritiidae (Acari: Oribatida) 1. Apotritia walkeri n.g., n.sp., from California. Acarologia, 33: 285-301.
Norton R.A., Sidorchuk E.A. 2014. Collohmannia johnstoni n. sp. (Acari, Oribatida) from West Virginia (U.S.A.), including description of ontogeny, setal variation, notes on biology and systematics of Collohmanniidae. Acarologia, 54: 271-334. doi:10.1051/acarologia/20142134 ![]()
Norton R.A., Behan-Pelletier V.M., Wang H.F. 1996a. The aquatic oribatid mite genus Mucronothrus in Canada and the Western USA (Acari, Trhypochthoniidae).- Canadian Journal of Zoology, 74: 926-949. doi:10.1139/z96-106 ![]()
Norton R.A., Graham T.B., Alberti G. 1996b. A rotifer-eating ameronothroid (Acari: Ameronothridae) mite from ephemeral pools on the Colorado Plateau. In: Mitchell R., Horn D.J., Needham G.R., Welbourn C.W. (Eds). Acarology IX – Proceedings. Volume 1. Columbus, Ohio: Ohio Biological Survey. p. 539-542.
Norton R.A., Alberti G., Weigmann G., Woas S. 1997. Porose integumental organs of oribatid mites (Acari, Oribatida). 1. Overview of types and distribution. Zoologica, Stuttgart, 146: 1-31.
Norton R.A., Franklin E., Crossley D.A. jr. 2010. Scapheremaeus rodickae n. sp. (Acari: Oribatida: Cymbaeremaeidae) associated with temporary rock pools in Georgia, with key to Scapheremaeus species in eastern USA and Canada. Zootaxa, 2393: 1-16.
Pachl P., Lindl A.C., Krause A., Scheu S., Schaefer I., Maraun M. 2017. The tropics as ancient cradle of oribatid mite diversity. Acarologia, 57: 309-322. doi:10.1051/acarologia/20164148 ![]()
Pauly F. 1956. Zur Biologie einiger Belbiden (Oribatei: Moosmilben) und zur Funktion ihrer pseudostigmatischen Organe. Zoologische Jahrbücher, Abteilung für Systematik, 84: 275-328.
Pfingstl T. 2013. Habitat use, feeding and reproductive traits of rocky-shore intertidal mites from Bermuda (Oribatida: Fortuyniidae and Selenoribatidae). Acarologia, 53: 369-382. doi:10.1051/acarologia/20132101 ![]()
Pfingstl T. 2015a. Morphological diversity in Selenoribates (Acari, Oribatida): new species from coasts of the Red Sea and the Indo-Pacific. International Journal of Acarology, 41: 356-370. doi:10.1080/01647954.2015.1035321 ![]()
Pfingstl T. 2015b. The intertidal Fortuyniidae (Acari, Oribatida): new species, morphological diversity, ecology and biogeography. Zootaxa, 3957: 351-382. doi:10.11646/zootaxa.3957.4.1 ![]()
Pfingstl T. 2017. The marine-associated lifestyle of ameronothroid mites (Acari, Oribatida) and its evolutionary origin: a review. Acarologia 57: 693-721.
Pfingstl T., Krisper G. 2011a. Juvenile stages of the arboricolous mite Cymbaeremaeus cymba (Nicolet, 1855) (Acari: Oribatida: Cymbaeremaeidae). International Journal of Acarology, 37: 175-189. doi:10.1080/01647954.2010.499373 ![]()
Pfingstl T., Krisper G. 2011b. The nymphs of Micreremus brevipes (Acari: Oribatida) and complementary remarks on the adult. Acta Zoologica Academiae Scientiarum Hungaricae, 57: 351-367.
Pfingstl T., Krisper G. 2014 Plastron respiration in marine intertidal oribatid mites (Acari, Fortuyniidae and Selenoribatidae). Zoomorphology, 133: 3359-378. doi:10.1007/s00435-014-0228-5 ![]()
Pfingstl T., Schuster R. 2012. Carinozetes nov. gen. (Acari: Oribatida) from Bermuda and remarks on the present status of the family Selenoribatidae. Acarologia, 52: 377-409. doi:10.1051/acarologia/20122067 ![]()
Pfingstl T., Schäffer S., Ebermann E., Krisper G. 2009a. Differentiation between two epilittoral species, Scutovertex arenocolus spec. nov. and Scutovertex pilosetosus Polderman (Acari: Oribatida) from different European coasts. Zootaxa, 2153: 35-54.
Pfingstl T., Schäffer S., Krisper G. 2009b. Morphological analysis of the juvenile stages of Provertex kuehnelti Mihelčič, 1959 (Acari, Oribatida). Acta Zoologica Academiae Scientiarum Hungaricae 55: 365-379
Pfingstl T., Schäffer S., Ebermann E., Krisper G. 2010a. The discovery of Scutovertex ianus sp. nov. (Acari, Oribatida) – a combined approach of comparative morphology, morphometry and molecular data. Contributions to Zoology, 79: 39-55.
Pfingstl T., Schäffer S., Krisper G. 2010b. Re-evaluation of the synonymy of Latovertex Mahunka, 1987 and Exochocepheus Woolley and Higgins, 1968 (Acari: Oribatida: Scutoverticidae). International Journal of Acarology, 36: 327-342. doi:10.1080/01647951003691988 ![]()
Pugh P.J.A., King P.E., Fordy M.R. 1987. A comparison of the structure and function of the cerotegument in two species of Cryptostigmata (Acari). Journal of Natural History, 21: 603-616. doi:10.1080/00222938700770371 ![]()
Schäffer S., Krisper G. 2007. Morphological analysis of the adult and juvenile instars of Scutovertex minutus (Acari, Oribatida, Scutoverticidae. Revue Suisse de Zoologie, 114: 663-683. doi:10.5962/bhl.part.80407 ![]()
Schäffer S., Köblmüller S., Pfingstl T., Sturmbauer C., Krisper G. 2010. Ancestral state reconstruction reveals multiple independent evolution of diagnostic morphological characters in the "Higher Oribatida" (Acari), conflicting with current classification schemes. BMC Evolutionary Biology, 10:246. [online http://www.biomedcentral.com/1471-2148/10/246 ![]() ] doi:10.1186/1471-2148-10-246
] doi:10.1186/1471-2148-10-246 ![]()
Schatz H., Behan-Pelletier V.M., OConnor B.M., Norton R.A. 2011. Suborder Oribatida van der Hammen, 1968. In: Zhang Z.-Q. (Ed.) Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa, 3148. p. 141-148.
Schubart H. 1975. Morphologische Grundlagen für die Klärung der Verwandtschaftsbeziehungen innerhalb der Milbenfamilie Ameronothridae (Acari). Zoologica, 123: 23-91.
Schulte G., Schuster R., Schubart H. 1975. Zur Verbreitung und Ökologie der Ameronothriden (Acari, Oribatei) in terrestrischen, limnischen und marinen Lebensräumen. Veröffentlichungen des Institutes für Meeresforschung Bremerhaven, 15: 359-385.
Sidorchuk E.A., Norton R.A. 2010. Redescription of the fossil oribatid mite Scutoribates perornatus, with implications for systematics of Unduloribatidae (Acari: Oribatida). Zootaxa, 2666: 45-67.
Smrž J. 2002. Microanatomical and microbiological characteristics of the quiescent state of Scutovertex minutus (Acari: Oribatida). Experimental and Applied Acarology, 27: 103-112. doi:10.1023/A:1021527904766 ![]()
Strenzke K. 1952. Untersuchungen über die Tiergemeinschaften des Bodens: Die Oribatiden und ihre Synusien in den Böden Norddeutschlands. Zoologica, Stuttgart, 104: 1-173.
Subías L.S. 2004. Listado sistemático, sinonímico y biogeográfico de los Ácaros Oribátidos (Acariformes, Oribatida) del mundo (1758-2002). Graellsia, 60, 3-305. doi:10.3989/graellsia.2004.v60.iExtra.218 ![]()
Suzuki K. 1978. Chaetotaxies of Japanese oribatids (2) Tectocepheus velatus. Atypus, 71: 51-56. (in Japanese).
Travé J. 1959. Dimorphisme sexuel chez Pirnodus detectidens Grandjean (Acariens, Oribates). Notes écologiques et éthologiques. Vie et Milieu, 9: 454-468.
Travé J. 1963a. Ecologie et biologie des Oribates (Acariens) saxicoles et arboricoles. Vie et Milieu, Supplement, 14: 1-267.
Travé J. 1963b. Oribates (Acariens) des Pyrenées-Orientales (1re serie). Provertex delamarei n. sp. Vie et Milieu, 13: 785-801.
Travé J., Fernandez N.A. 1986. Contribution a la connaissance du genre Scapheremaeus: S. argentinensis, n. sp. (Oribate). Acarologia, 27: 349-359.
Travé J., Vachon M. 1975. François Grandjean 1882-1975 (Notice biographique et bibliographique). Acarologia, 17: 1-19.
Travé J., André H.M., Taberly G., Bernini F. 1996. Les Acariens Oribates. Wavre, Belgium: AGAR & Société internationale des Acarologues de Langue française. pp. 110.
Vanschoenwinkel B., De Vries C., Seaman M., Brendonck L. 2007. The role of metacommunity processes in shaping invertebrate rock pool communities along a dispersal gradients. Oikos, 116: 1255-1266. doi:10.1111/j.0030-1299.2007.15860.x ![]()
Vanschoenwinkel B., Gielen S., Seaman M., Brendonck L. 2008a. Any way the wind blows – frequent wind dispersal drives species sorting in ephemeral aquatic communities. Oikos, 117: 125-134. doi:10.1111/j.2007.0030-1299.16349.x ![]()
Vanschoenwinkel B., Gielen S., Vandewaerde H., Seaman M., Brendonck L. 2008b. Relative importance of different dispersal vectors for small aquatic invertebrates in a rock pool metacommunity. Ecography, 31: 567-577. doi:10.1111/j.0906-7590.2008.05442.x ![]()
Vanschoenwinkel B., Gielen S., Seaman M., Brendonck L. 2009. Wind mediated dispersal of freshwater invertebrates in a rock pool metacommunity: Differences in dispersal capacities and modes. Hydrobiologia, 635: 363-372. doi:10.1007/s10750-009-9929-z ![]()
Vitzthum H. 1943. Acarina. In: Bronn H.G. (Ed.). Klassen und Ordnungen des Tierreiches, 5. Band: Arthropoda, IV. Abteilung: Arachnoidea, 5. Buch: Acarina. Leipzig: Becker & Erler Kom.-Ges. p. 1011.
Wallwork J.A. 1962. Notes on the genus Pertorgunia Dalenius, 1958 from Antarctica and Macquarie (Acari: Oribatei). Pacific Insects, 4: 881-885.
Wallwork J.A. 1964. A revision of the family Podacaridae Grandj. (Acari: Oribatei). Acarologia, 6: 387-399.
Wallwork J.A. 1967. Cryptostigmata (oribatid mites). In: Gressit J.L. (Ed.). Entomology of Antarctica. Antarctic Research Series, vol. 10. Washington, D.C: American Geophysical Union. pp. 105-122. doi:10.1029/AR010p0105 ![]()
Wallwork J.A. 1981. A new aquatic oribatid mite from Western Australia (Acari: Cryptostigmata: Ameronothridae). Acarologia, 22: 333-339.
Walter D.E. 1995. Dancing on the head of a pin: mites in the rainforest canopy. Records of the Western Australian Museum, Supplement, 52: 49-53.
Walter D.E. 1999. Living on leaves: Adaptations of Australian rainforest mites. In: Needham G.R., Mitchell R., Horn D.J., Welbourn C.W. (Eds). Acarology IX. Volume 2, Symposia. Columbus, Ohio: Ohio Biological Survey. pp. 73-78.
Walter D.E., Behan-Pelletier V.M. 1993. Systematics and ecology of Adhaesozetes polyphyllos sp.nov. (Acari: Oribatida: Licneremaeoidea), a leaf inhabiting mite from Australian rainforests. Canadian Journal of Zoology, 71: 1024-1040. doi:10.1139/z93-136 ![]()
Weigmann G. 2006. Hornmilben (Oribatida). Die Tierwelt Deutschlands. Teil 76. Keltern: Goecke & Evers. pp. 520.
Weigmann G. 2010. Reconstruction of stem species pattern as a strategy towards integrated phylogenetic systematics and taxonomy, applied to early-derivative Poronota (Oribatida).Acarologia, 50: 291-315. doi:10.1051/acarologia/20101972 ![]()
Weigmann G., Schulte G. 1977. The evolution of the family Ameronothridae (Acari: Oribatei). I. Morphological aspects. Acarologia, 19: 161-166.
Willmann C. 1931. Moosmilben oder Oribatiden (Cryptostigmata). In: Dahl F. (Ed.). Die Tierwelt Deutschlands, Band 22, vol. 5. Jena: Gustav Fischer. p. 79-200.
Woas S. 1990. Die phylogenetischen Entwicklungslininen der Höheren Oribatiden (Acari) I. Zur Monophylie der Poronota Grandjean, 1953. Andrias, Karlsruhe, 7: 91-168.
Woas S. 1991. Distribution of characters and phylogenetic age – systematic problems in the higher taxa of the Oribatida. In: Schuster R., Murphy P.W. (Eds). The Acari – Reproduction, development and life-history strategies. London, New York: Chapman and Hall. pp. 329-334. doi:10.1007/978-94-011-3102-5_26 ![]()
Woas S. 1992. Beitrag zur Revision der Gymnodamaeidae Grandjean, 1954 (Acari, Oribatei). Andrias, Karlsruhe, 9: 121-161.
Woas S. 1998. Mosaikverteilung der Merkmale basaler Höherer Oribatiden – Die Gattungen Passalozetes und Scutovertex. In: Ebermann E. (Ed.). Arthropod Biology: Contributions to Morphology, Ecology and Systematics. Biosystematics and Ecology Series, 14: 291-313.
Woas S. 2000. Die Gattungen Poroliodes und Cymbaeremaeus und ihr verwandtschaftliches Umfeld. Carolinea, Karlsruhe, 58: 165-181.
Woas S. 2002. Acari: Oribatida. In: Adis J. (Ed.). Amazonian Arachnida and Myriapoda. Sofia, Moscow: Pensoft Publishers. pp. 21-291.
Xue X.F., Dong Y., Deng W., Hong X.Y., Shao R. 2017. The phylogenetic position of eriophyoid mites (superfamily Eriophyoidea) in Acariformes inferred from the sequences of mitochondrial genomes and nuclear small subunit (18S) rRNA gene. Molecular Phylogenetics and Evolution, 109: 271-282. doi:10.1016/j.ympev.2017.01.009 ![]()
acarologia_4258_supplementaryvideo.mp4



2017-12-12
Date accepted:
2018-02-21
Date published:
2018-06-01
Edited by:
Sidorchuk, Ekaterina

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Norton, Roy A. and Franklin, Elizabeth
 Download article Download low definition
Download article Download low definitionDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)