




Morphological ontogeny, distribution of Hermannia scabra (Acari: Oribatida: Hermanniidae) in Svalbard and descriptive population parameters
Seniczak, Stanisław1 ; Seniczak, Anna2 and Coulson, Stephen J.3
1✉ Department of Evolutionary Biology, Faculty of Natural Sciences, Kazimierz Wielki Uni-versity, Ossoli ´nskich 12, Pl 85-092 Bydgoszcz, Poland.
2Department of Ecology, UTP University of Science and Technology, Kordeckiego 20, Pl 85-225 Bydgoszcz, Poland.
3Department of Arctic Biology, University Centre in Svalbard, P.O. Box 156, 9171 Long-yearbyen, Norway and ArtDatabanken The Swedish Species Information Centre, Swedish University of Agricul-tural Sciences, Box 7007, 750 07 Uppsala, Sweden.
2017 - Volume: 57 Issue: 4 pages: 877-892
https://doi.org/10.24349/acarologia/20174214ZooBank LSID: 8A0EC9C9-3441-4991-917E-383B1E6BD337
Keywords
Abstract
Hermannia Nicolet, 1855 (Hermanniidae) is included in the cohort Nothrina (Norton and Behan-Pelletier 2009) but due to the brachypyline venter of the adult it is also treated as an intermediate genus between the lower (macropyline) and higher (brachypyline) oribatid mites (Norton and Behan-Pelletier 2009; Colloff 2011; Seniczak et al. 2017). Seniczak et al. (2017) compared the morphological ontogeny of three species of Hermannia with two species of Phyllhermannia Berlese, 1916 and 16 other species of Crotonioidea and stated that the latter group is highly differentiated and insufficiently known, and therefore it is difficult to determine the correct systematic position of Hermannia in this superfamily. For example, Hermannia retains seta f 1, which is ancestral in Nothrina, but loses the second pair of exobothridial seta (exi), which is progressive in early-derivative oribatid mites. Hermannia also retains seta f 2 in all instars, as most species of Brachypylina, but the ontogeny of leg setae of Hermannia species more closely resembles the macropyline than the brachypyline oribatid mites. Subías (2017) listed in Hermannia sensu stricto a total of 17 species, but six of them he considered inquirenda.
The diagnostic characters of Hermannia species are based exclusively on the morphology of adults, mainly the forms of the bothridial seta and prodorsal seta in, the ornamentation of the cuticle, and the form of the setae on the notogaster and legs (Sitnikova 1975; Woas 1978, 1981; Weigmann 2006). The body length of adults of most Hermannia species overlaps and only have a diagnostic value in comparing the small and large species. The nymphs are strikingly similar to adults, except for the smaller body size and folded and less sclerotized cuticle (Bäumler 1970; Travé 1977a; Ermilov et al. 2012; Seniczak et al. 2017).
The juveniles of Hermannia are inadequately known. According to Norton and Ermilov (2014) and Seniczak et al. (2017), the full morphological ontogeny of only three species is studied:
(1) Hermannia gibba (C. L. Koch, 1839): Nicolet (1855) described and illustrated a nymph, Grandjean (1933) studied the formulae of gastronotal, genital, aggenital and anal regions, and line of dehiscence and hysterosomal setation (Grandjean 1947) in a wide phylogenetic context. Bäumler (1970) investigated the larva and nymphs, and Ermilov et al. (2012) described and illustrated the morphological ontogeny of juveniles, including leg setae and solenidia.
(2) Hermannia jesti Travé, 1977: Travé (1977a) described all juvenile stages and numerical ontogeny of leg setae, illustrated the tritonymph and leg I of the larva, and studied epimeral setae in a wide phylogenetic context (Travé 1977b).
(3) Hermannia reticulata Thorell, 1871: Thor (1930) gave photos of a nymph in ventral and lateral aspects, Grandjean (1949) studied the formulae of gastronotal, genital, aggenital and anal regions of this species and Seniczak et al. (2017) described and illustrated the morphological ontogeny, including leg setae.
The known morphological ontogeny of some other Hermannia species is incomplete. According to Norton and Ermilov (2014), Oudemans (1900) and Willmann (1931) described and illustrated a nymph of Hermannia convexa (C. L. Koch, 1839), and Hammen (1978) described and illustrated the ventral aspect of all juvenile stages of this species, including the prelarva, and discussed the morphology and development of chelicera in a wide phylogenetic context. However, in our opinion a nymph described and illustrated by Willmann (1931) belongs to Hermannia scabra (L. Koch, 1879) because it has similar body shape and the pattern of the cuticle on the gastronotum is as in the nymphs investigated here. However, the description of a nymph by Willmann (1931) is general and omits important diagnostic characters. We therefore redescibe this stage and describe the morphology of the other juvenile instars that are currently undescribed. Hermannia scabra is a large species, similarly to congener H. reticulata (Woas 1978), but was rarely recorded from Svalbard (Coulson and Refseth 2004; Coulson 2008; Bayartogtokh et al. 2011), its presence in Svalbard was even uncertain (Coulson et al. 2014); its dis-tribution and ecology in this archipelago is poorly known.
The aim of this paper is to describe the morphological ontogeny of H. scabra, expand the diagnosis of this species to include morphological characters of the juveniles and compare its ontogeny with that of congeners. We also present the distribution of H. scabra in Svalbard, population density, and stage structure of this species.
We investigated the distribution of H. scabra in the Svalbard archipelago (Figure 1A) using 174 samples from 33 locations (Table 1, Figure 1B). Samples were collected by S. J. Coulson during the polar summer, in a period of around six weeks (from late June to early August) between 2009 – 2014. Soil samples were collected in the typical vegetation communities at each location, and the number of samples from each location varied between 2 – 12. Each sample had dimensions of 10 x 10 cm by ca. five cm deep (the usual maximum depth of the organic soil). The samples (soil and plant cover) were kept cool until being extracted in Tullgren funnels (Burkard Scientific Ltd., Uxbridge, UK) at the University Centre in Svalbard (UNIS) within five days from collection and preserved in 96% ethanol. We determined the density, stage structure and the body length using all mites, but in the abundant two samples (Kapp Lee b, Petuniabukta a) we measured 15 individuals selected randomly.




Statistical calculations were done on the body length of H. scabra in vegetation classes 8 and 16, where this species was abundant. Normality of data was examined by the Kolmogorov-Smirnov test and the homogeneity of variance with the Levene’s test. To find differences between the body length we used ANOVA variance analysis (Stanisz 2006). The level of significance for all statistical tests was accepted at α = 0.05. Data were analyzed using MS Excel 2007, Statistica 12.0.
The juveniles and adults of H. scabra used in the morphological study were collected from vegetation class 8 (wet vegetated flats, beaches, slopes and river fans with some exposed Dryas vegetation and graminoids Luzula sp.) in Svalbard (Figure 1B). The ontogeny of this species is similar to that of H. reticulata (Seniczak et al. 2017), so the illustrations of H. scabra are limited to the body regions that show substantial differences. Illustrations were prepared from individuals temporarily mounted on slides in lactic acid. In the description of instars and figures we used the following abbreviations: prodorsal setae (ro, le, in, ex), bothridium (bo), bothridial seta (bs), notogastral or gastronotal setae (c-, d-, l-, h-, p-series), adanal and anal setae (ad-, an-series), aggenital seta (ag), epimeral setae (1a–c, 2a, 3a–c, 4a–c), cupules or lyrifissures (ih, ips, iad), opisthonotal gland opening (gla), leg solenidia (Σ, Φ, Ω), famulus ($pepsilon$) and setae (bv, d, ft, tc, it, p, u, a, s, pv, pl, v). In H. scabra, the hypertrichy of epimeral setae occurs, and the hypertrichous setae were labelled as a' or b'. Terminology follows that of Grandjean (1933, 1947, 1949, 1953) and Norton and Behan-Pelletier (2009). The species nomenclature follows that of Norton and Ermilov (2014).
We sampled oribatid mites in the vegetation classes and locations according to a revised vegetation map of the Svalbard (Johansen et al. 2012). Of the total 11 investigated vegetation classes, H. reticulata was absent from seven (classes 9, 10, 12, 14, 15, 17 and 18), but was present in four:
(1) Class 8 (wet vegetated flats, beaches, slopes and river fans), which is represented by Kapp Lee. This pioneer vegetation also consists of exposed Dryas vegetation and graminoids (Luzula sp.).
(2) Class 11 (moss tundra), such as Kapp Laila and occurs in areas with moderate snow cover during winter. Most often developed in the lower parts of hill slopes, on established river fans and in small depressions. The vegetation is a mixture of Cassiope tetragona D. Donand and moderate snowbed species, including Salix polaris Wahlenb., Dryas octopetala L., Equisetum arvense L., Saxifraga oppositifolia L. and Silene acaulis (L.) Jacq. The moss layer is generally moderately developed.
(3) Class 13 (Arctic meadows – rich vegetation), present in five regions (Fjortendejulibukta, Fuglehuken, Reindyrflya, Storholmen, and Signehamn). This class consists of luxuriant vegetation communities characterized by grasses and forbs with a high species diversity. These communities are associated with warm south and southwest facing slopes with available water during the growing season. To some extent the luxuriant bird cliff vegetation, established densely vegetated river fans and the drier parts of Dupontia meadows are included in this map unit.
(4) Class 16 (established dense Dryas heaths), which is exemplified by six regions (Kvadehuken, Mosselbukta, Petuniabukta, Ringhorndalen, Sørkapp and Tjuvfjord), with a more heavy snow cover dominated by D. octopetala. The most common variant is poor in species, while others may contain herbs, sedges and lichens. In depressions and less drained sites mosses in combination with C. tetragona are developed, whereas on coastal plains S. oppositifolia partly with lichens form the dominant cover. In mountain areas and in most of the northern and eastern regions of the archipelago Dryas stands with Papaver dahlianum (Tolm.) Elven & Nilsson are common.
Nothrus scaber L. Koch, 1879.
Hermannia (Heterohermannia) scabra: Woas 1992; Subías 2004, 2017.
Hermannia scabra: Woas 1978; Mehl 1979; Karppinen and Krivolutsky 1982; Golosova et al. 1983; Schatz 1983; Tarman 1983; Marshall et al. 1987; Bernini et al. 1995; Olszanowski et al. 1996.
Morphology of adult — Adult large (815 – 1,050 μm, Table 2), stocky and brown, with characters of Hermannia as given by Woas (1978). Our individuals smaller (815 – 945 μm, n= 30, Table 3) than those investigated by Woas (1978) (1,000 – 1,150 μm), with notogastral setae relatively longer (Figure 2A) than in Woas. In our adults, number of epimeral setae larger (3-1-4/5-6/8, most common formula 3-1-5-7, Table 4) than in Woas (3-1-5-5/6). In our adults two pairs of large aggenital setae occur (Figure 3A), whereas in Woas five pairs of small setae present. In our individuals additional anal seta rarely occurred asymmetrically on one anal plate. Leg segments as in H. reticulata (Seniczak et al. 2017) and relatively thicker than in Woas, but leg setae similar. Coupled seta d present at leg solenidia Σ, Φ and Φ1 (Table 5). Formula of setae (+solenidia) of legs (trochanter to tarsus) of our adults: I – 1-5-(5+1)- (5+2)- (21+2), II – 1-5-(5+1)-(5+1)-(14+2); III – 2-3-(4+1)-(4+1)-13; IV – 1-3-4-(4+1)-13.





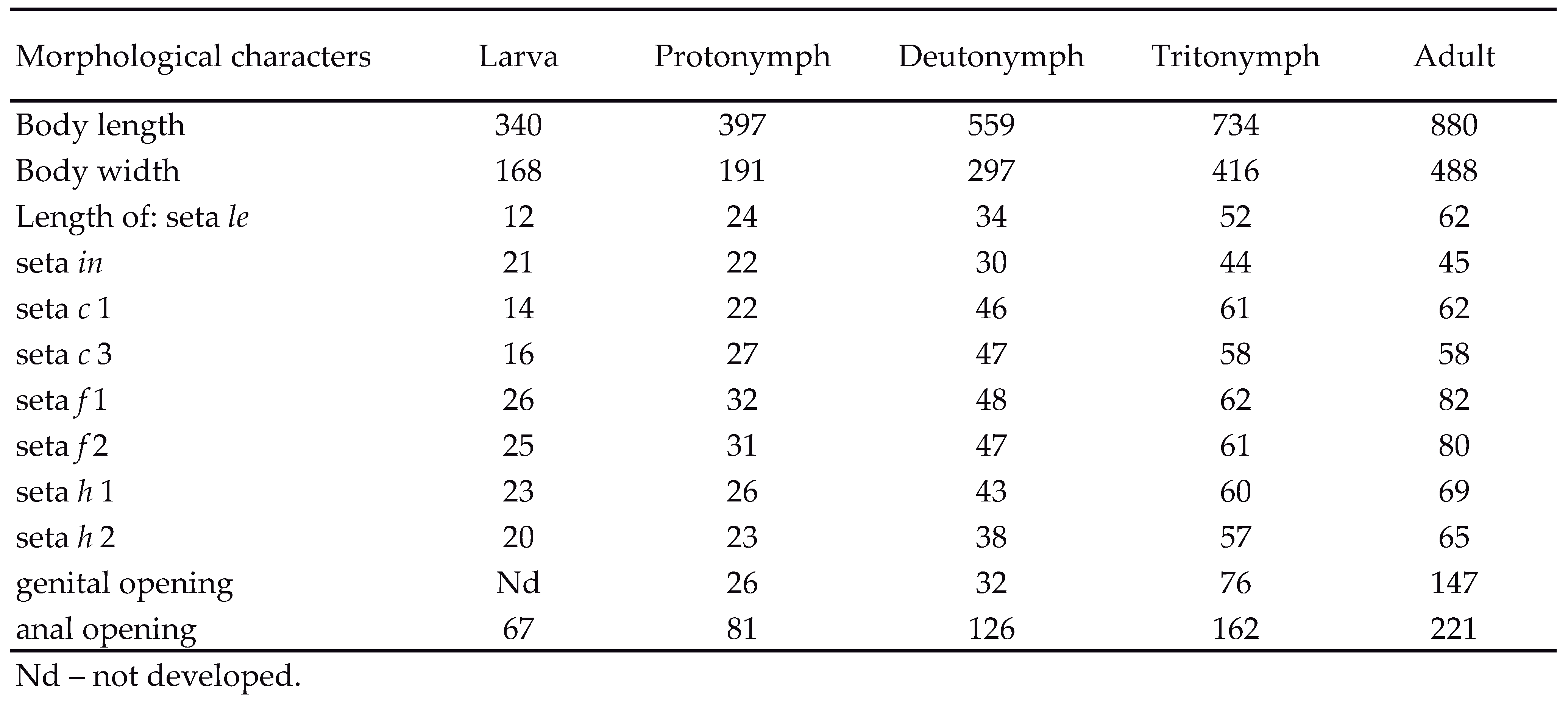


Diagnosis of juveniles — Juveniles stocky, oval in dorsal aspect and light brown, most prodorsal and gastronotal setae of medium size and phylliform. Anterior and medial part of prodorsum reticulate, bothridial seta clavate. Gastronotum uneven, most setae on apophyses. Formulae of epimeral setae: 3-1-2 (larva), 3-1-2-2 (protonymph), 3-1-3/4-4/5 (deutonymph) and 3-1-3/4-5/6 (tritonymph). Most setae on femora, genua and tibiae phylliform.
Description of juvenile stages — Larva stocky, oval in dorsal aspect and light brown, prodorsum subtriangular. Rostrum rounded (Figure 2D), prodorsal setae ro and le short, seta in slightly longer (Table 2), all phyliform (Figure 2G); mutual distance of pair in more than three times that of pairs le and ro. Opening of bothridium small, rounded (Figure 2F), with minute seta ex inserted close to lateral part of bothridial opening, bothridial seta short, clavate, head with short and thick barbs. Anterior part of prodorsum reticulate (Figure 2E), other parts folded (Figure 2G).
Gastronotum of larva with 12 pairs of setae, including h 2, positioned lateral to medial part of anal opening, h 3 absent (Figures 2H, 3B). All gastronotal setae of medium size (Table 2), phylliform, length slightly increasing from anterior to posterior. Gastronotum uneven, most gastronotal setae on large apophyses. Paraproctal valves (segment PS) glabrous. Cupules not evident in folded integument, except for ih located lateral to anterior part of anal opening (Figure 3B), opisthonotal gland opening (gla) anteromedial to seta f 2 (Figures 2D, H). Line of dehiscence visible in shape of inverted letter Y. Formula of epimeral setae: 3-1-2, all setae short, except for scaliform 1c covering Claparède’s organ. Number of leg setae and solenidia as in H. reticulata (Seniczak et al. 2017), but most setae of femora, genua and tibia phylliform.
Prodorsum, prodorsal setae, bothridium and bothridial seta of protonymph as in larva, but prodorsal reticulation of cuticle denser than in larva and hysterosoma widest along 2/3 of its length. Gastronotum of protonymph uneven and with 16 pairs of setae due to appearance of setae h 3 and p-series (Figure 3C) that remain in other nymphs (Figures 3D, E); most setae on apophyses, setae h 1 and h 2 on large posterior apophysis. In protonymph gastronotal setae of medium size and phyliform; length of setae slightly increasing from anterior to posterior. In protonymph one pair of genital setae appears (Figure 3C), and three pairs are added in deutonymph and tritonymph each (Figures 3D, E); all of medium size and smooth. In protonymph, two pairs of setae appear on epimere IV, 4a on unsclerotized integument with slating striae, and 4b on sclerotized epimere IV (Figure 3C). In deutonymph and tritonymph one or two setae added on epimere III and IV each (Figures 3D, E), including 4a' located on unsclerotized integument in deutonymph (if present unilaterally) and tritonymph; all short. Ontogeny of epimeral setae shown in Figures 4A–D. In deutonymph one pair of long aggenital setae appear, and three pairs of short adanal setae, in tritonymph second pair of long aggenital setae appears and two pairs of anal setae. In deutonymph and tritonymph, setae h 2 and h 3 on large posterior apophysis (Figures 3D, E, 2J). Paraproctal valves of protonymph and deutonymph glabrous, tritonymph with two pairs of short anal setae (Figure 3E). Seta d present at base of leg solenidia Σ, Φ and Φ1, most setae on femora, genua and tibiae phyl-liform, and distribution of setae and solenidia as in H. reticulata (Seniczak et al. 2017), but ontogeny of setae differs (Tables 5, 6).


Summary of ontogenetic transformations — Prodorsal and gastronotal setae remain short or medium sized in all instars, seta ex is tiny, located on the lateral edge of bothridial opening, and the bothridial seta remains clavate. The larva has 12 pairs of gastronotal setae, and the nymphs and adult have 16 pairs. The epimeral formulae of H. scabra are: 3-1-2 (larva), 3-1-2-2 (protonymph), 3-1-3/4-4/5 (deutonymph), 3-1-3/4-5/6 (tritonymph) and 3-1-4/5-6/8 (adult). The number of epimeral setae varies in adults between locations in Svalbard, especially on epimere IV, and the most common formula is 3-1-5-7 (Table 4). Seta 4a' occupies the posterolateral position to seta 4a in the deutonymphs (if present), tritonymph and adult, whereas other epimeral setae are inserted in anterior row. The formula of genital setae is 1-4-7-9 (protonymph to adult), aggenital setae is 1-2-2 (deutonymph to adult), gastronotal setae is 12-16-16-16-16 and the formula of segments PS–AN is 03333-0333-022. The shape of leg solenidia and most setae remains similar in all instars, but the number of setae increases during ontogeny (Table 5), mainly in the protonymph and deutonymph.




Distribution, ecology and biology — Hermannia scabra has a holarctic distribution (Subías 2004, 2017; Bayartogtokh et al. 2011). This species is an unspecialized panphytophage (Schatz 1983; Honciuc and Lundqvist 2009) and prefers moist and wet localities (Hull 1916; Trägårdh 1931; Sellnick 1949; Hammen 1952) with moss and algae, especially on coastal rocks, in salt marshes (Gjelstrup and Solhøy 1994), and dead plant material or fungi (Brodo 2000).
Hull (1916) considers H. scabra common in maritime localities, semiaquatic habitat and on dead wood, whereas Trägårdh (1931) found this species under stones and on driftwood on the sea shore and in freshwater algae. Hammer (1969) noted H. scabra on lily bulbs and in soil ferns with Sansevieria sp. and various plants and roots, whereas Behan and Hill (1978) found this species in moss and tussock tundra. Purvis (1982) found H. scabra relatively abundant in undecomposed gorse needles (Ulex europaeus L.) in a coastal shrub of southeast Ireland, and on the earthen banks with granite boulders to a height about 1 m, whereas Żbikowska-Zdun et al. (2006) found this species abundant in moss and lichens covering an old 18th-century gate in the Bug River Protected Landscape Area in southern Poland. This species was also found on the feathers of black kite (Milvus migrans Boddaert) and sparrow hawk (Accipiter nisus L.) (Krivolutsky and Lebedeva 2004) and in bird collecting sites, where also the juveniles of this mite species were present (Lebedeva and Lebedev 2007).
This species was cultured by Jalil (1965) in the laboratory (20°C) on decaying skeletonized leaves collected from broadleaved woodland. The whole development of species lasted 184 days, the development time of subsequent stages increasing during the ontogeny (egg 15 days, larva 16 days, protonymph 41 days, deutonymph 42 days and tritonymph 70 days). This author also observed that the development of H. scabra in the field, under oak Quercus robur L. in Sutton Bonnington (England, Canada), required a full year. He observed oviposition of H. scabra from September to November with most larvae in October. The greatest total density of this species was in October, with numbers declining during the winter and was lowest from May to July when the larvae were absent.
We found H. scabra in four of the 11 vegetation classes, at four of the 33 locations sampled (Figure 2, Table 1), and in seven of the 174 samples. Constancy of this spe-cies in this archipelago was therefore very low (4%). If we consider only the locations occupied by H. scabra, the greatest constancy of this species was at Kapp Lee (100%) and the lowest was at Petuniabukta (8%). The greatest mean density of H. scabra (11,800 individuals per 1 m2) was noted in vegetation class 8 at Kapp Lee (wet vegetated flats, beaches, slopes and river fans with some exposed Dryas vegetation and graminoids Luzula sp.), but the density varied here between 200 – 45800 individuals per 1 m2. A lower density was observed in vegetation class 16 (established dense Dryas heaths) at Petuniabukta (15,000 individuals per 1 m2), whereas in other vegetation classes the density was very low (Table 1).
The juveniles of H. scabra dominated the adults, comprising 89% of all individuals (Table 3), but the proportion of juveniles in the population varied between 88 – 100%. Among the juveniles, the protonymphs dominated and the tritonymphs were least abundant. The stage structure of H. scabra slightly varied among samples. For example, in the most abundant population at Kapp Lee, the juveniles comprised 91% of all individuals, and the stage structure of this species was 73 larvae, 204 protonymphs, 129 deutonymphs, 9 tritonymphs and 43 adults, whereas in the less abundant population of this species at Petuniabukta, the juveniles formed 89% of all individuals and the stage structure was 3 larvae, 78 protonymphs, 20 deutonymphs, 32 tritonymphs and 17 adults.
The sex ratio of H. scabra was not determined because of the deep brown pigmentation and ornamentation of adults which made it impossible to determine sex by transmission light microscopy. No eggs were observed inside females. The body length of the adults in vegetation class 8 was significantly higher than in vegetation class 16, but generally it varied greatly within sampling locations, largest and smallest individuals both occurring at Kapp Lee, where the mean density of this species was the highest.
We compare the morphological ontogeny of H. scabra studied here with that of H. gibba, H. jesti and H. reticulata, studied by Ermilov et al. (2012), Travé (1977a) and Seniczak et al. (2017) respectively. These species differ from one another by the following important morphological characters:
(1) Shape of the bothridial seta in all instars: in H. reticulata and H. scabra this seta is clavate, whereas in other species it is setiform.
(2) Pattern of cuticle on the prodorsum of larva: in H. scabra the cuticle is reticulate, whereas in other species the pattern is indistinct or unknown.
(3) Pattern of cuticle on the prodorsum of nymphs: in H. reticulata and H. scabra the cuticle is reticulate, whereas in other species the pattern is indistinct.
(4) These species also differ from each other by some other morphological characters and the ontogeny of epimeral setae (Table 7).



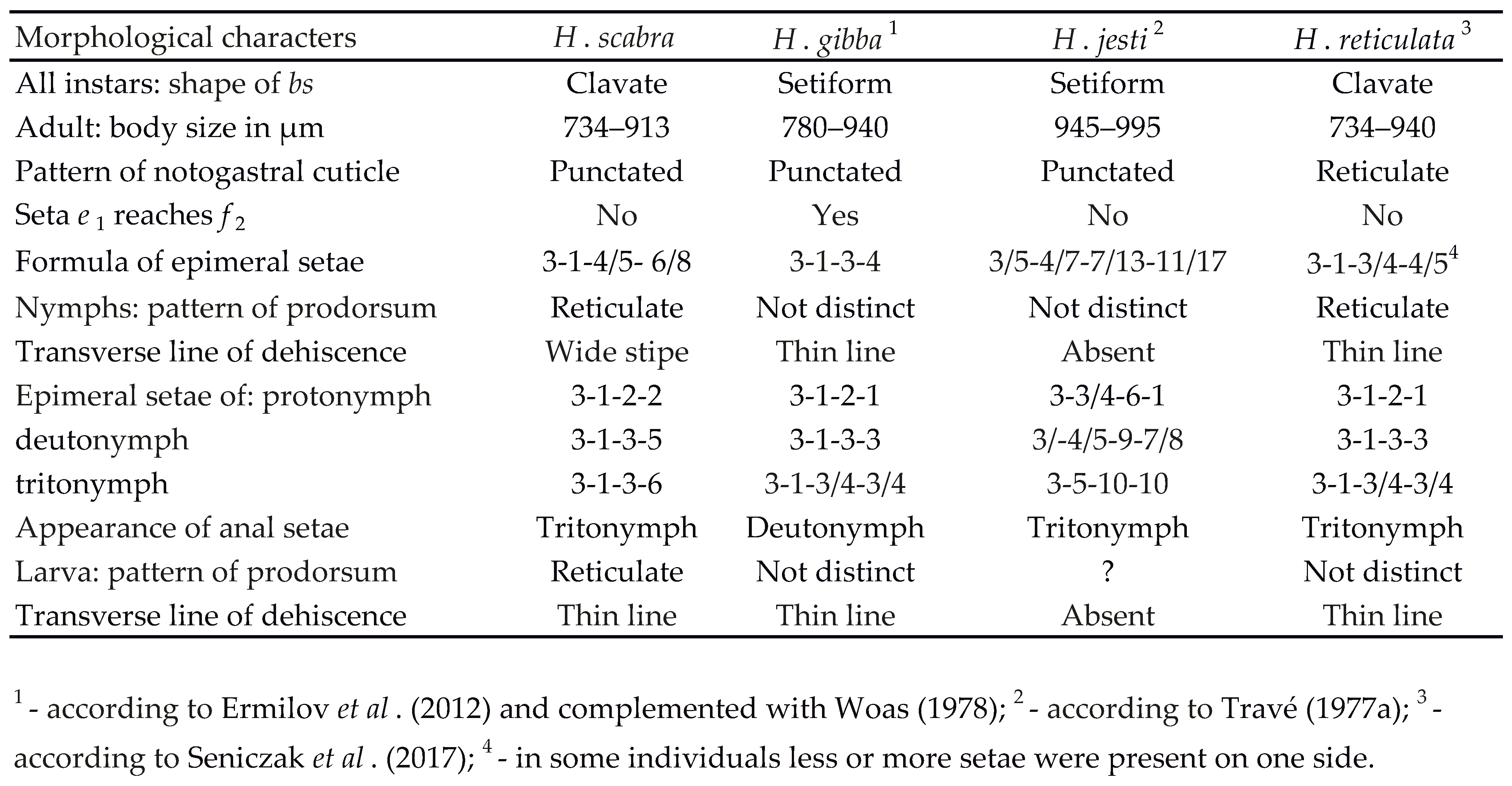
This comparison shows the great similarity of juvenile H. reticulata and H. scabra, which differ from one another mainly by the pattern of cuticle on the prodorsum and gastronotum and number of epimeral setae. These species have a clavate bothridial seta, whereas other species have it setiform. Interestingly, H. scabra differs from all other species by the presence of two setae on epimere IV of protonymph, instead of one in the other species. One of these setae (4a) is inserted on unsclerotized integument and occupies this position in deutonymphs and tritonymph. In deutonymph, one of the new setae (4a') is also is inserted on unsclerotized integument, posterolateral to seta 4a, and occupies this position in tritonymph and adult. Particularly characteristic is the ontogeny of leg setae, which differs in H. gibba, H. reticulata and H. scabra (Table 6). Travé (1977a) gave the numerical ontogeny of leg setae of H. jesti, but it differs from that of H. gibba, H. reticulata and H. scabra. For example, femur I of protonymph of H. jesti gains one seta, comparing to the larva, whereas other species gain more setae. Thus, each of four species has its own ontogeny of these setae (Table 6).
The nymphs of H. scabra investigated here are similar to a nymph described and illustrated by Willmann (1931); both have similar pattern of cuticle of gastronotum and a large posterior apophysis bearing setae of h-series. This author observed similarity of morphology of this nymph to that of Camisia.
The body length of the adult of H. scabra described by L. Koch (1879) from Novaja Zemlja (Russia) was 1,000 μm, whereas the individuals from Svalbard studied here were slightly smaller (815 – 945 μm) and similar to those studied by Willmann (1931) (length 900 μm). The maximal body lengths of this species given by Sitnikova (1975) and Woas (1978) are larger (1,550 μm and 1,150 μm, respectively), but all authors noted that the notogaster of H. scabra is thickly punctated as in our individuals.
We found H. scabra investigated here to be identical to those studied by Willmann (1931), both as adult and nymphs. In Willmann (1931) the adult of H. scabra has six pairs of notogastral setae in inner row (c 1, d 1, e 1, f 1, h 1 and p 1), and seta p 2 is located relatively far from p 1, as in our adults, whereas in Sitnikova (1975) and Woas (1978) seta p 2 is located close to p 1. Moreover, the adult of H. scabra investigated by Woas (1978) has relatively shorter notogastral setae than in our individuals and has five short pairs, whereas our adults have two pairs of long setae.
The juveniles of H. scabra add new morphological characters to our knowledge on the morphology of Hermannia species, such as a new formula of epi-meral setae and new distribution of these setae on epimere IV, new pattern of ontogeny of leg setae, and the presence of a wide transverse line of dehiscence on the gastronotum of nymphs, which in other species of Hermannia is expressed only by a narrow line (Grandjean 1947; Ermilov et al. 2012; Seniczak et al. 2017). The line of dehiscence is also observed in the juvenile instars of Phyllhermannia (Colloff 2011), in the nymphs of C. lapponica and in all juveniles of C. voleolata (Seniczak 1991).
The ontogeny of epimeral setae of H. scabra is interesting because two pairs of setae appear on epimere IV of protonymph, instead of one pair in other Hermannia species, and the inner setae (1a, 2a, 3a, 4a, 4a') are inserted on unsclerotized integument between epimeres, which is unique in Hermannia; in other species of this genus all setae are on epimeres. In this species the hypertrichy occurs on epimeres III and IV, as in H. jesti and H. reticulata (Travé 1977a; Seniczak et al. 2017). Grandjean (1934) observed in Nanhermannia nana (Nicolet, 1855) the appearance of seta 4a in the protonymph, seta 4b in the deutonymph (anteromedial to seta 4a, as in most species of Brachypylina), and setae 4c and 4d in the tritonymph (lateral to 4a), and all remain in the adult. However, the ontogeny of epimeral setae of H. scabra is different because two setae (4a, 4b) appear in the protonymph, one seta (4c) in the deutonymph and hypertrichy in 3- and 4-rows occurs in the deutonymph, tritonymph and adult. In Nothrus species, the hypertrichy of epimeral setae appears mainly on epimere I (Seniczak 1992; Seniczak and Żelazna 1992; Seniczak and Norton 1993).
Hypertrichy makes the setal notation problematic. Probably this is why many authors omitted notation of epimeral setae in Hermannia (Bäumler 1970; Travé 1977a, b; Woas 1978; Olszanowski 1996; Ermilov et al. 2012). Colloff (2011) labelled these setae in the closely related genus Phyllhermannia, where hypertrichy occurs mainly on epimere IV of the tritonymph and adult. Generally, Colloff's setal labeling is positional, not indicating the homologies. The exception is Phyllhermannia sauli Colloff, 2011, in which his seta 4b appears on epimere IV of protonymph, and two setae are added in the deutonymph, including his 4a that is added medial to 4b, as in Nanhermannia (Grandjean 1934). In Ph. lemannae Colloff, 2011, setal labeling is positional, but his seta 4a is also added medial to 4b, as in Ph. sauli.
The protonymph of H. reticulata also has one seta on epimere IV (4a), but two setae (4b, 4c) are added laterally to 4a during the ontogeny (Seniczak et al. 2017). In this species, the hypertrichy rarely occurs in the tritonymph, but is common in the adult. In contrast, the protonymph of H. scabra has two setae (4a, 4b) on epimere IV, and seta 4c is added laterally to 4b in the deutonymph. In the latter stage the hypertrichy occurs, and develops in the tritonymph and adult. In both species, the hypertrichous setae are added in unpredictable positions and numbers on each epimere, and normal setae can also be absent, which makes labeling of epimeral setae difficult.
In the tritonymph of H. gibba, H. reticulata and H. scabra, the most common formula of epimeral setae of is 3-1-3-3 (Ermilov et al. 2012; Seniczak et al. 2017), which suggests that three letters (a, b, c) are sufficient to label the normal epimeral setae in this genus, whereas other setae (a', b') are hypertrichous. However, in Nanhermannia, a shift of the deutonymphal seta 4b to the epimere III occurs (Grandjean 1934), whereas in Hermannia or Phyllhermannia it does not because the apodemes 3 and 4 are long, almost transverse, and the epimeral setae of 3- and 4-series are in their own rows.
Holotrichous setation of nymphs is not common in Crotonioidea, it occurs in Hermannia, Phyllhermannia and Nothrus C.L. Koch, 1836; other genera lose on the gastronotum at least seta f 1. However, the nymphs of Hermannia differ from those of other genera by the body form, ornamentation of the body and other morphological characters given by Seniczak et al. (2017).
Adult size variation in the Svalbard population was small and the largest individuals were only 16% longer than the smallest. However, the body size of H. scabra varies dramatically between geographically distinct populations (Willmann 1931; Sitnikova 1975; Woas 1978, 900 – 1,550 μm), which can pose the question concerning the identity of this species in this body range. Moreover, the number of aggenital setae of H. scabra varies greatly. For example, Woas (1978) noted five pairs of aggenital setae in the adults, but in those investigated here only two pairs are present, as in H. reticulata and H. jesti (Seniczak et al. 2017; Travé 1977a). In H. gigantea Sitnikova, 1977, there are also five pairs of aggenital setae present, and the body length (1,193 – 1,211 μm) includes that of H. scabra, and the notogaster is thickly punctated. All these data encourage further study of H. scabra from different geographical regions to determine the variability of the body length and aggenital setation of this species or define the cryptic species that may exist.
Hermannia scabra is a typical component of oribatid mite communities inhabiting the Arctic ecosystems in the Svalbard archipelago. It is not as common in this archipelago as H. reticulata, which has a higher constancy (12%, Seniczak et al. 2017) than that of H. scabra (4%). However, H. scabra forms larger agglomerations and its maximal density is greater (45,800 individuals per 1 m2) than that of H. reticulata (10,300 individuals per 1 m2), due to more abundant juvenile stages. In four vegetation classes we also observed more individuals of the former species (624 individuals) than the latter species (425 individuals).
Using the example of H. scabra and H. reticulata, it is clear how the distribution of species in the Arctic ecosystems depends on ecology of species. These species do not occur together because the former species prefers the moist and wet localities (Trägårdh 1931; Hammen 1952), such as vegetation class 8 (wet vegetated flats, beaches, slopes and river fans with some exposed Dryas vegetation and graminoids Luzula sp.) and class 16 (established dense Dryas heaths), where H. reticulata is absent (Seniczak et al. 2017). In contrast, H. reticulata prefers dry habitats, like heaths, fell field, meadows, moss and lichens (Behan and Hill 1978), the chamaephyte vegetation (Hammer 1944, 1946) and lichens (Gjel-strup 1978), and in Svalbard archipelago it mainly occupies vegetation class 12 (rich moss tundra, bird cliff vegetation) where H. scabra is absent. Both species occur in vegetation class 16, but H. scabra was abundant in one sample in Petuniabukta, where H. reticulata was absent, whereas H. reticulata occurred in one sample at Mosselbukta and in one sample at Sørkapp (Seniczak et al. 2014), where H. scabra was absent. Interestingly, the body length of the adults of H. scabra in vegetation class 8 significantly exceeded that in vegetation class 16, whereas in H. reticulata such differences were not observed (Seniczak et al. 2017).
Generally, the proportion of H. scabra juveniles was distinctly higher (89 – 100% of total mites) than that of H. reticulata (52 – 61%). In other Arctic oribatid species as Mycobates sarekensis (Trägårdh, 1910) and Oribatella arctica (Thor, 1930), the juveniles constituted 36% and nearly 5% of populations, respectively (Seniczak et al. 2015a, b). The distribution of H. scabra in Svalbard indicates that the abundant agglomerations of this species in vegetation classes 8 and 16 are important sources of its expansion into surrounding areas using, for example passive transport by birds (Krivolutsky and Lebedeva 2004) or other animals. In several samples in these, and other vegetation classes, the juveniles and adults of this species were present or only adults or juveniles, which may indicate the development of H. scabra in the adjacent habitats.
We thank Dr. H. Tømmervik for kindly providing the vegetation information for the locations at which H. scabra was collected from, the anonymous reviewers for helpful suggestions, and the subject editor of Acarologia, Dr. Ekaterina A. Sidorchuk for fruitful discussion on the ontogeny of epimeral setae.
Bayartogtokh B., Schatz H., Ekrem T. 2011 — Distribution and diversity of the soil mites of Svalbard, with redescriptions of three known species (Acari: Oribatida) — Intern. J. Acar., 37: 467-484. doi:10.1080/01647954.2010.525525 ![]()
Bäumler W. 1970 — On the morphology, biology and ecology of Hermannia gibba (C.L. Koch) (Acarina: Oribatei) taking into account some accompanying species — Z. angew. Entomol., 66: 337-362.
Behan V.M., Hill S.B. 1978 — Feeding habits and spore dispersal of oribatid mites in the North American Arctic — Rev. Ecol. Biol. Sol, 15: 497-516.
Berlese A. 1916: — Centuria prima di Acari nuovi — Redia, 12: 19-67.
Bernini F., Castagnoli M., Nannelli R. 1995 — Arachnida, Acari. In: Minelli A., Rufo S., La Posta S. (eds.). Checklist delle specie della fauna italiana, 24 — Bologna: Calderini, p. 1-131.
Brodo F. 2000 — The insects, mites, and spiders of Hot Weather Creek, Ellesmere Island, Nunavut. In: Gameau M., Alt B.T. (eds.): Environmental Response to Climate Change in the Canadian High Arctic — Geolog. Survey of Canada, Bull., 529: 145-173.
Colloff M.J. 2011 — New species of the oribatid mite genus Phyllhermannia Berlese, 1916 (Acari, Oribatida, Hermanniidae) from wet forests in south-eastern Australia show a high diversity of morphologically-similar, short-range endemics — Zootaxa, 2770: 1-60.
Coulson S.J. 2008 — The terrestrial invertebrate fauna of Svalbard: a cross referenced check-list. ( http://www.unis.no/35_STAFF/staff_webpages/biology/steve_coulson/ docu-ments/Fullreport_008.pdf ![]() ) (actualized in 15 May 2008).
) (actualized in 15 May 2008).
Coulson S.J., Refseth D. 2004 — The terrestrial and freshwater invertebrate fauna of Svalbard (and Jan Mayen). In: Prestrud P., Strøm H., Goldman H. (eds.). A catalogue of the terrestrial and marine animals of Svalbard. Skrifter 201 — Norwegian Polar Institute, Tromsø; p. 57-122.
Coulson S.J., Convey P., Aakra K., Aarvik L., Ávila-Jiménez M.L., Babenko A., Biersma E.M., Boström S., Brittain J.E., Carlsson A.M., Christoffersen K., De Smet W.H., Ekremj T., Fjellberg A., Füreder L., Gustafssonm D., Gwiazdowicz D.J., Hansen L.O., Holmstrup M., Hullé M., Kaczmarek Ł., Kolicka M., Kuklin V., Lakka H.K., Lebedeva N., Makarova O., Maraldo K., Melekhina E., Ødegaard F., Pilskog H.E., Simon J..C, Sohlenius B., Solhøy T., Søli G., Stur E., Tanasevitch A., Taskaeva A., Velle G., Zawierucha K., Zmud-czyńska-Skarbek K. 2014 — The terrestrial and freshwater invertebrate biodiversity of the archipelagoes of the Barents Sea; Svalbard, Franz Josef Land and Novaya Zemlya — Soil Biol. Biochem., 68: 440-470. doi:10.1016/j.soilbio.2013.10.006 ![]()
Ermilov S.G., Ryabinin N.A., Anichkin A.E. 2012 — Morphology of juvenile instars of two oribatid mite species of the family Hermanniidae (Acari, Oribatida) — Zool. Zhurn., 91: 657-668. doi:10.1134/S0013873812070081 ![]()
Gjelstrup P. 1978 — Oribatid mites (Acarina) from the Faroe Islands — Norw. J. Entomol., 25(1): 45-50.
Gjelstrup P., Solhøy T. 1994 — The oribatid mites (Acari) of Iceland. The Zoology of Iceland, Steenstrupia — Zool. Mus. Copenhagen, 3, 57e: 1-78.
Golosova L.D., Karppinen E., Krivolutsky D.A. 1983 — List of oribatid mites (Acarina, Oribatei) of northern Palaearctic region. II. Siberia and the Far East — Acta Entomol. Fenn., 43: 1-14.
Grandjean F. 1933 — Étude sur le développement des Oribates — Bull. Soc. zool. France, 58: 30-61.
Grandjean F. 1934 — Les poils des épimères chez les Oribates (Acariens) — Bull. Mus. nat. Hist. natur. (2), 6: 504-512.
Grandjean F. 1947 — Observations sur les Oribates. (18e série) — Bull. Mus. Nat. Hist. Nat., 2e série, 19: 395-402.
Grandjean F. 1949 — Formules anales, gastronotiques, génitales et aggénitales du développement numérique des poils chez les Oribates — Bull. Soc. zool. France, 74: 201-225.
Grandjean F. 1953 — Essai de classification des Oribates (Acariens) — Bull. Soc. zool. France, 78: 421-446.
Hammen L., van der. 1952 — The Oribatei (Acari) of the Netherlands — Zool. Verh., Leiden, 17: 1-139.
Hammen L., van der. 1978 — The evolution of the chelicerate life-cycle — Acta Biotheoretica, 27: 44–60. doi:10.1007/BF00048403 ![]()
Hammer M. 1944 — Studies on the oribatids and Collemboles of Greenland — Medd. Grönland, Copenhagen, 141(3): 1-210.
Hammer M. 1946 — Oribatids. In: The Zoology of East Greenland — Medd. Grönland, Copenhagen, 122(1): 1-39.
Hammer M. 1969 — Oribatids found at plant quarantine stations in U.S.A. — Vid. med. Dansk naturh. foren., 132: 63–78.
Honciuc V., Lundqvist L. 2009 — Diversity and species distribution of oribatid mites (Acari-Oribatida) in a geographical and ecological unique area of southern Sweden — Romanian J. Biology – Zoology, 54(1) :7-17. http://lup.lub.lu.se/record/948593 ![]()
Hull J.E. 1916 — Terrestrial Acari of the Tyne Province, I. Oribatidae — Trans. Nat. Hist. soc. Northumberl., n. s., 4: 381-410.
Jalil M. 1965 — The life cycle of Hermannia scabra (L. Koch 1879) (Acarina – Oribatei) — Oikos, 16: 6-19. doi:10.2307/3564860 ![]()
Johansen B.E., Karlsen S.R., Tømmervik H. 2012 — Vegetation mapping of Svalbard utilising Landsat TM/ETM+ data — Polar Record, 48 (244): 47-63. doi:10.1017/S0032247411000647 ![]()
Karppinen E., Krivolutsky D.A. 1982 — List of Oribatid mites (Acarina, Oribatei) of northern Palaearctic region. I. Europe — Acta Entomol. Fenn., 41: 1-18.
Koch C.L. 1836 — Deutschlands Crustaceen, Myriapoden und Arachniden — Regensburg, Bd. 4-9.
Koch C.L. 1839 — Deutschlands Crustaceen, Myriapoden und Arachniden — Regensburg, Heft 28-30.
Koch L. 1879 — Arachniden aus Sibirien und Novaja Zemlja, eingesammelt von der Schwedischen Expedition im Jahre 1875 — Kongl. Svenska Vet. Handl., Stockholm, 16(5): 1-136.
Krivolutsky D.A., Lebedeva N.V. 2004 — Oribatid mites (Oribatei, Acariformes) in bird feathers: nonpasserines — Acta Zool. Lithuanica, 14: 26-47.
Lebedeva N.V., Lebedev V.D. 2007 — Diversity of oribatid mites (Acari, Oribatei) and other soil microarthropods in plumage of raptors — Caucasian Entomol. Bull., 3: 9-18.
Marshall V.G., Reeves R.M., Norton R.A. 1987 — Catalogue of the Oribatida (Acari) of Continental United States and Canada — Mem. Entomol. Soc. Canada, 139: 1-418. doi:10.4039/entm119139fv ![]()
Mehl R. 1979 — Checklist of Norwegian ticks and mites (Acari) — Fauna Norvegica, B, 26: 1-45.
Nicolet H. 1855 — Histoire naturelle des Acariens qui se trouvent aux environs de Paris — Arch. Mus. Hist. Nat. Paris, 7: 381-482.
Norton R.A., Behan-Pelletier V.M. 2009 — Suborder Oribatida. In: Krantz G.W., Walter D.E. (eds.). A Manual of Acarology — Texas Tech Univ. Press, Lubbock: 430-564. doi:10.1653/024.092.0323 ![]()
Norton R.A., Ermilov S.G. 2014 — Catalogue and historical overview of juvenile instars of oribatid mites (Acari: Oribatida) — Zootaxa, 3833: 1-132. doi:10.11646/zootaxa.3833.1.1 ![]()
Olszanowski Z. 1996 — A monograph of the Nothridae and Camisiidae of Poland (Acari: Oribatida: Crotonioidea) — Genus, Supplement, Wrocław; p. 1–201.
Olszanowski Z., Rajski A., Niedbała W. 1996 — Roztocze Acari – Mechowce Oribatida. Katalog Fauny Polski – Catalogus faunae poloniae — Sorus, Poznań, 34(9): 1-243.
Oudemans A.C. 1900 — Further notes on Acari — Tijdschr. Entomol., 43: 109-128.
Purvis G. 1982 — The soil arthropod fauna (Acari and Collembola) of a coastal locality in southeast Ireland — J. Life Sci. Royal Dublin Society, 3: 379-396.
Sellnick M. 1949 — Milben von der Küste Schwedens — Entomol. tidskr., 70: 123-135.
Schatz H. 1983 — U.-Ordn.: Oribatei, Hornmilben — Catalogus Faunae Austriae, Wien, Teil IXi: 1-118.
Seniczak S. 1991 — The morphology of juvenile stages of moss mites of the family Camisiidae (Acari: Oribatida), VI — Zool. Anz., 227: 331-342.
Seniczak S. 1992 — The morphology of juvenile stages of moss mites of the family Nothridae (Acari: Oribatida). I — Zool. Anz., 229: 134-148.
Seniczak S., Norton R.A. 1993 — The morphology of juvenile stages of moss mites of the family Nothridae (Acari: Oribatida). III — Zool. Anz., 230: 19-33.
Seniczak S., Seniczak A., Coulson S.J. 2015a — Morphology, distribution and biology of Mycobates sarekensis (Acari: Oribatida: Punctoribatidae) — Intern. J. Acar., 41: 663-675. doi:10.1080/01647954.2015.1102968 ![]()
Seniczak S., Seniczak A., Coulson S.J. 2015b — Morphology, distribution and certain population parameters of the Arctic mite Oribatella arctica (Acari: Oribatida: Oribatellidae) — Intern. J. Acar., 41: 395-414. doi:10.1080/01647954.2015.1048727 ![]()
Seniczak S., Seniczak A., Gwiazdowicz D.J., Coulson S.J. 2014 — Community structure of oribatid and gamasid mites (Acari) in moss-grass tundra in Svalbard (Spitsbergen, Norway) — Arctic Ant. Alpine Res., 46: 591-599. doi:10.1657/1938-4246-46.3.591 ![]()
Seniczak S., Seniczak A., Coulson S.J. 2017 — Morphological ontogeny, distribution, and descriptive population parameters of Hermannia reticulata (Acari: Oribatida: Hermanniidae), with comments on Crotonioidea — Intern. J. Acar., 43(1): 52-72. doi:10.1080/01647954.2016.1229812 ![]()
Seniczak S., Żelazna E. 1992 — The morphology of juvenile stages of moss mites of the family Nothridae (Acari: Oribatida). II — Zool. Anz., 229: 149-162.
Sitnikova G.L. 1975 — Family Hermanniidae Sellnick, 1928. In: Ghilarov M.S., editor. Key to soil-inhabiting mites – Sarcoptiformes — Nauka Publisher, Moscow: 101-104. [In Russian]
Sitnikova L.G. 1977 — New species of of the oribatid families Ameronothridae and Hermanniidae (Oribatei) forthe fauna of the USSR — Parazit. Sb., 27: 160-168. [In Russian]
Stanisz A. 2006 — Easy course of statistic using Statistica PL and medicine examples. 1. Basic statistic — StatSoft Polska, Kraków. (In Polish)
Subías L.S. 2004, 2017 — Listado sistemático, sinonímico y biogeográfico de los Ácaros Oribátidos (Acariformes, Oribatida) del mundo (1758-2002) — Graellsia, 60: 3-305. [Actualized in February 2017: 1-598]. Available from: ( http://bba.bioucm.es/cont/docs/RO_1.pdf ![]() ). doi:10.3989/graellsia.2004.v60.iExtra.218
). doi:10.3989/graellsia.2004.v60.iExtra.218 ![]()
Tarman K. 1983 — Catalogus faunae Jugoslaviae III/4 Acarina Oribatei. Consilium Academiarum Scientiarum Rei Publicae Socialistae Foederativae Jugoslaviae — Acad. Sci. Artium Slovenica, Ljubljana, 3: 1-61.
Thor S. 1930 — Beiträge zur Kenntnis der Invertebratenfauna von Svalbard — Skr. Svalb. Ishavet, Oslo, 27: 1-156.
Thorell T. 1871 — Om Arachnider fran Spitzbergen och Beeren-Eiland — Öfvers. Kongl. Vet. Akad. Förh., 28: 683-702.
Travé J. 1977a — Hermannia jesti Travé (Oribatida, Hermanniidae), Oribate du Népal — Acarologia, 19: 697-710.
Travé J. 1977b — La néotrichie épimérique d'Hermannia jesti sp. n. (Oribate) — Acarologia, 19: 123-131.
Trägardh I. 1910 — Acariden aus dem Sarekgebirge — Naturw. Unters. Sarek., Zool., Stockholm, 4: 375-586.
Trägårdh I. 1931— Terrestrial Acarina — Zoology of the Faroes, Copenhagen, 49: 1-69.
Weigmann G. 2006 — Hornmilben (Oribatida). In: Dahl F., series founder. Die Tierwelt Deutschlands, part 76 — Keltern, Goecke & Evers: 1-520.
Willmann C. 1931 — Moosmilben oder Oribatiden (Cryptostigmata). In: Dahl F., series founder. Die Tierwelt Deutschlands — Gustav Fischer Jena, 22: 79-200.
Woas S. 1978 — Die Arten der Gattung Hermannia Nicolet 1855 (Acari, Oribatei) I — Beitr. naturk. Forsch. Südw. Deutschl., 37: 113-141.
Woas S. 1981 — Zur Taxonomie und Phylogenie der Hermanniidae Sellnick 1928 (Acari, Oribatei) — Andrias, Karlsruhe, 1: 7-88.
Woas S. 1992 — Hermannia (Phylhermannia) neotropica, eine neue Hermanniidae aus Südbrasilien (Acari: Oribatei) — Andrias, Karlsruhe, 9: 163-178.
Żbikowska-Zdun K., Piksa K., Watrak I. 2006 — Diversity of mites (Acari: Oribatida) in selected microhabitats of the Bug river protected landscape area — Biol. Lett., Poznań, 43: 277-286.



2017-02-07
Date accepted:
2017-05-02
Date published:
2017-10-10
Edited by:
Sidorchuk, Ekaterina

This work is licensed under a Creative Commons Attribution 4.0 International License
2017 Seniczak, Stanisław; Seniczak, Anna and Coulson, Stephen J.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)