




Ants as predators of the Spinose Ear Tick, Otobius megnini (Dugès) in Sri Lanka
Diyes, G.C.P.1 ; Karunarathna, N.B.2 ; Silva, T.H.S.E.3 ; Karunaratne, W.A.I.P.4 and Rajakaruna, R.S.5
1Department of Zoology, University of Peradeniya, Peradeniya, 20400, Sri Lanka
2Department of Zoology, University of Peradeniya, Peradeniya, 20400, Sri Lanka
3Department of Zoology, University of Peradeniya, Peradeniya, 20400, Sri Lanka
4Department of Zoology, University of Peradeniya, Peradeniya, 20400, Sri Lanka
5✉ Department of Zoology, University of Peradeniya, Peradeniya, 20400, Sri Lanka
2017 - Volume: 57 Issue: 4 pages: 747-753
https://doi.org/10.24349/acarologia/20174200Keywords
Abstract
Tick infestations are very hard to control owing to the wide distribution of certain species, their remarkable longevity, seasonal dynamics and off-host development (Sonenshine, 1993). The most widely used and currently effective method is the use of chemical acaricides like carbamate, organophosphate, synthetic pyrethroid, formamidine, macrocyclic lactone and pyrazole (Lovis et al., 2011). Although effective, intensive use can result in the development of resistance (Foil et al., 2004; Guerrero et al., 2012), impaired environmental and human health with negative effects on non-target organisms and poor quality animal products (e.g., milk, meat and hide; Rajput et al., 2006). Other control methods such as vaccination (Willadsen, 1995), selection of resistant host breeds (Samish et al., 2004; Shyma et al., 2015) and pasture spelling which involves rotating livestock among paddocks for periods of 3-4 months to starve the larval ticks (Wilkinson, 1957) have been practiced, particularly for controlling the cattle tick, Rhipicephalus microplus. Application of acaricide substitutes such as extracts of plants like Azadirachta indica, Calotropis procera, Nicotiana tabacum (Zaman et al., 2012) are being promoted. Moreover, in Sri Lanka, local remedies are widely being used among the rural people to remove or to repel ticks from livestock and pets. Some of these methods include: use of coconut oil, citronella oil, neem (Azadirachta indica) oil/leaves/seeds, marigold plant leaves and use of mothballs (Personal communications with villagers and livestock farmers).
Numerous potential biological control agents of ticks including pathogens, parasitoids and predators have been documented (Jenkins 1964; Mwangi and Kaaya 1997; Samish and Rehacek, 1999; Kaaya, 2003). Pathogens like bacteria, fungi and nematodes that infect and kill ticks, parasitoids that deposit their eggs in ticks or predators like birds and ants have been suggested as potential candidates for controlling some hard and soft tick species under field and laboratory conditions (Samish et al., 2004). Predator-tick relationships of 28 arthropod families have been identified, of which, many are ants (Hymenoptera: Formicidae), followed by carabid beetles (Coleoptera: Carabidae) and some spiders (Araneae: Lycosidae; Samish and Alekseev, 2001). Among ants, Aphaenogaster, Formica, Iridomyrmex, Monomorium, Notoncus, Pheidole, Pogonomyrmex, Rhytidoponera and Solenopsis are some of the genera that occasionally target ticks as their food source (Samish and Rehacek, 1999). Ants that feed on ticks are considered as bio-suppressors (Samish et al., 2004). They are generalist predators that feed on ticks occasionally and may affect the size of a tick population in nature, but using them as biological control agents may have negative consequences as it requires a large increase in the ant population which could lead to changes in non-target species (Symondson et al., 2002) or result in the ant becoming a pest (Barbosa, 1998; Bellows and Fisher, 1999). Gleim et al. (2013) discussed the potential use of Solenopsis invicta against tick species: Amblyomma americanum and Amblyomma maculatum populations in burned habitats. However, few studies have examined the exact effect of ants on tick population dynamics. Hence, the importance of ants in biological control is still controversial (Barbosa, 1998).
The spinose ear tick, Otobius megnini (Acari: Ixodida: Argasidae) is a soft tick of medical and veterinary importance that feeds in the ear canals of wide range of domesticated animals including horses (Diyes and Rajakaruna, 2016a), and occasionally humans, causing otoacariasis in Sri Lanka (Ariyarathne et al., 2016). When present in the ear canal, they can cause paralysis, irritations, toxic conditions, allergies, eardrum perforation, muscle spasms, severe otitis and act as vectors of Q fever (Jellison et al., 1948; Madigan et al., 1995; Estrada-Peña and Jongejan, 1999). Larvae and nymphs of the spinose ear tick are parasitic while adults (males and females) and unfed larvae are free-living and found in cracks and crevices in the immediate vicinity of the host (Sonenshine, 1993). Here we present the first report of predatory behavior of four genera of ants on O. megnini in Sri Lanka.
Larvae and nymphs of Otobius megnini infesting the ear canals of stabled horses at the Nuwara Eliya race course (GPS: 6.962829N, 80.769207E) were collected onto small pieces of white open-wove cotton bandage. The ground cover materials in stables were brushed carefully using a soft bristle duster to collect the free-living adults. Live ticks were brought to the Parasitology Laboratory in the Department of Zoology, University of Peradeniya and were counted, weighed and separated according to life stage. Some of the field-caught engorged nymphs were allowed to moult into adults. Field-caught adults were allowed to mate and lay eggs. Some of these eggs were kept in perforated Eppendorf® tubes (1.5 ml each containing 100 eggs) to obtain unfed larvae. The rest of the eggs were placed in perforated Eppendorf® tubes (1.5 ml each containing 100 eggs) and were kept on a microcentrifuge rack. Field caught engorged larvae and nymphs, and the laboratory moulted adults were placed in quadrangular plastic grid plates (25 wells in each plate and three ticks per well). The grid plates were covered with a plastic mesh which allowed ants to enter but excluded flies (e.g., the scuttle fly, Megaselia scalaris; Diyes et al., 2015) and other larger predators. Larvae and nymphs with fungal infections were removed from the collection. Eppendorf® tubes containing eggs/unfed larvae and the grid plates containing larvae, nymphs and adults were kept on a laboratory bench and observed for 72 h at three-hour intervals. All plates were opened at 0600 h and closed with the plastic lid at 1800 h. The whole set up was left on the bench near a window under laboratory conditions at 28±2 °C and 82% - 90% relative humidity. When ants were observed feeding on the ticks or eggs, these ants were preserved in absolute ethanol and the tick stage being fed on was noted. A few weeks after, the same set up was again placed on the bench near a window (preferably in a different location of the lab) with a new generation of ticks and a different local ant colony. This was repeated over a five-month period from September 2014 to January 2015. Ant species that fed on ticks were morphologically identified according to standard keys and descriptions (Bolton, 1994; Dias, 2002). In a second experiment, laboratory-hatched adult ticks (n=15 for each exposure) were placed in a sealed plastic container with sawdust to simulate its natural habitat in stables. Adults were found actively burrowing into the sawdust. Ant species that fed on adult ticks in the first set of experiments were introduced separately to the laboratory bench near the plastic container with the sawdust and adult ticks. The container was again covered with plastic mesh and kept on the laboratory bench to determine whether the ants could locate ticks at the bottom of the sawdust layer. Ants preying on ticks were observed every two days at three-hour intervals from 0600-1800 h each day. At each observation time, the adult ticks in the container were carefully observed for at least 5 mins. On the following day, all the containers were sealed and kept away from ants. Ants that actively attacked the ticks, either feeding or biting, were isolated and the physical damage done by them was noted. The feeding preference of the different ant species on each tick life stage was analyzed using a chi-square test.
A total of 500 eggs and 500 unfed larvae (5 replicate batches of 100 eggs and larvae), 575 engorged larvae, 650 engorged nymphs and 450 adults were exposed to ants. Ants preyed on eggs, larvae (both engorged and un-engorged) and adults (both males and females in the sawdust and in the grid plates), but not on nymphs. Five ant species belonging to four genera: Monomorium spp. Mayr, 1855 (small black ant), Pheidole sp. Westwood, 1839 (bigheaded ant), Tapinoma melanocephalum Fabricius, 1793 (ghost ant) and Crematogaster sp. Lund, 1831 (acrobat ant) were found predating the different tick life stages. A single ant species was found in each genus except in Monomorium where there were two species: Monomorium sp. 1 and Monomorium sp. 2 (Table 1). Overall, there was a significant difference in the predation preference of five ant species (Chi-square test Χ2=88.33, p<0.001; Table 1). Tapinoma melanocephalum which fed on eggs and adults was the best predator followed by Crematogaster sp. which fed on engorged larvae and adults while Monomorium sp. 2 which fed only on few unfed larvae and adults can be considered as the least distractive tick predator. Individual comparisons of the feeding preferences of five ant species showed significant differences for all species pairs (Chi-square test, p<0.05) except between Monomorium sp.1 and Pheidole sp. (Χ2=0.24, p=0.630).

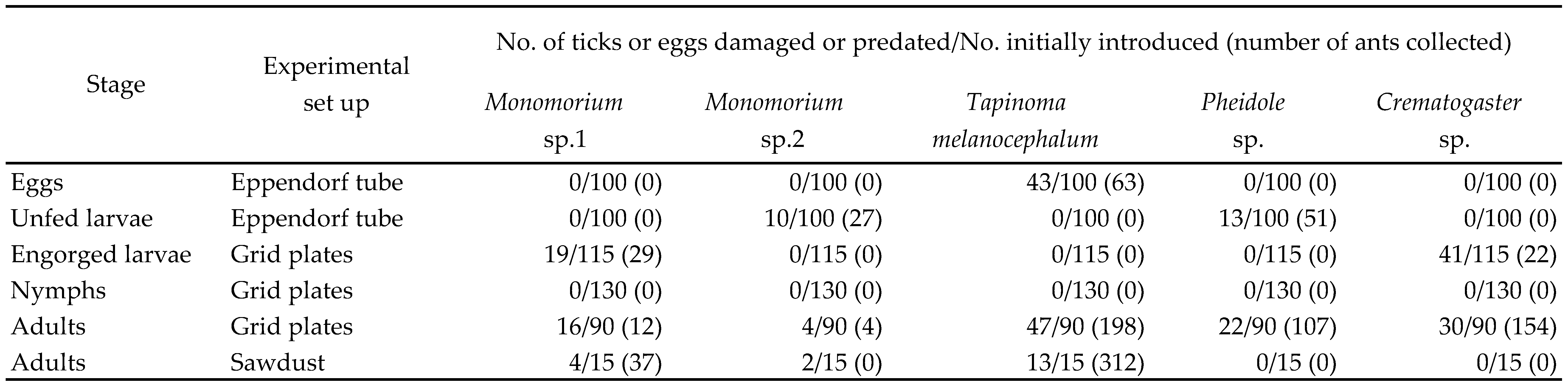
Among the five ant species, Monomorium sp. 1 predated on engorged larvae while Monomorium sp. 2 predated on unfed larvae (Table 1). Crematogaster sp. predated on engorged larvae and adults where some engorged larvae and adults were completely eaten and the rest were badly wounded. Tapinoma melanocephalum was the only ant species that fed on eggs of O. megnini in addition to adults and caused the highest damage to the tick population. Pheidole sp. attacked both unfed larvae and adults. There was a significant difference in the predatory preference of ant species for the different tick life stages (eggs, larvae and adults; Chi-square test, Χ2=112.2, p<0.001). Eggs were preferred over unfed larvae (Χ2=6.5, p=0.01) and adults were preferred over eggs and larvae (Χ2=112.2, p<0.001). There was no significant difference in the feeding fed larvae and unfed larvae (Χ2=1.0, p=0.308) or the adults in grid plates and adults in sawdust (Χ2=0.041, p=0.840).
Both species of Monomorium sp. and T. melanocephalum were the only ant species that actively burrowed in the sawdust to prey on adult ticks. To escape from the ants, some of the burrowed ticks came to the surface of the sawdust, most of which already had wounds from the ant attacks. The majority of ticks (87%) that came to the surface died due to T. melanocephalum attacks.
Parish (1949) observed that two ant species, the small black ant Monomorium minimum and the large red ant Pogonomyrmex barbatus var. molefaciens, preyed on nymphs and adults of O. megnini. Monomorium minimum has also been reported preying on other soft ticks such as Argas miniatus and A. persicus (Bishopp, 1913) and Solenopsis invicta was reported to prey on Ornithondoros moubata and O. parkeri (Oliver et al., 1986). Among ants, anti-tick activity has been described for the genus Pheidole. For example, Pheidole weiseri is known to attack Rhipicephalus microplus (Wilkinson, 1970) and P. megacephala is known to attack Amblyomma cajennense, Rhipicephalus appendiculatus and R. microplus in different parts of the world (Rodriguez et al., 1983; Castineiras et al., 1987). Furthermore, Samish and Rehacek (1999) recognized 27 such ant species of 16 genera as tick predators. However, information on the anti-tick activity of T. melanocephalum and Crematogaster sp. is scant. Yet, T. melanocephalum is reported as the primary predator of Rhodnius prolixus eggs, a vector of Chagas' disease (Gomez-Nunez, 1971) and some Crematogaster species are recognized as biological control agents for Leptopharsa gibbicarina which is known to induce pestalotiopsis on palm leaves (Montañez et al., 1998) and Dorylus quadratus which is known to attack honey bees (Adgaba et al., 2014).
The spinose ear tick is found in all purebred, stabled horses whose ear hairs have been shaven, but not in those with unshaven ear hair. These ticks are also not present in non-purebred horses or when horses are kept in open range (Diyes and Rajakaruna, 2016a). Seasonal tick abundance shows high larval counts during warmer months of the year due to high egg hatching rates (Diyes and Rajakaruna, 2016b). Tick infestation has become a serious problem for the stable managers of the Nuwara Eliya racecourse, as these horses are of significant commercial value for racing and other recreational purposes. Although many control measures have been attempted, controlling the free-living stages has proved difficult. To use ants as a control measure for O. megnini, ant species that feed on free living stages could be bred at the race course and released into the dumping yard of sawdust to establish local colonies. However, using ants as a biological control agent of O. megnini has its own limitations. For example, laboratory observations cannot be easily extrapolated to a natural context because ants may forage differently when they are in more variable field conditions (Gromadzki and Bull, 1997). Moreover, anti-tick activity may greatly depend on environmental factors like time of the day, brightness, humidity, geographical area and season of the year (Gromadzki and Bull, 1997; Sutherst and Maywald, 2005). Further, ant density may change with tick population dynamics and this need to be monitored in order to understand the long-term existence of ant colonies and their specific predator prey interactions. For ants to be a successful biological control agent, they should have long hunting seasons, large populations of workers that cover a large area, and should be nonspecific with regard to prey life stage (Fisher et al., 1999).
Since ants are considered as generalist predators that feed occasionally on ticks, ant populations do not likely depend on the size of tick populations to persist (Samish et al., 2004). Symondson et al. (2002) discussed that generalist predators can sometimes affect the size of tick populations in nature, but manipulating these predator populations to reduce tick numbers may require large increases in predator population sizes, which could also cause changes in other ant populations present in the natural ecosystem. Others argue that the use of ants as a biological control agent is unfeasible because they will have negative impacts on non-target organisms including humans or livestock and may later require secondary control measures (Holway et al., 2002). Moreover, sometimes ants unintentionally help the dispersion of ticks as ants carry tick eggs back to the colony (eg. extitT. melanocephalum). Therefore, more attention has to be paid to the use of ants as tick predators to determine their effectiveness in biological control and the consequences. The ant genera which were observed predating O. megnini during the present study are common with a wide distribution in Sri Lanka (Dias, 2002) and are therefore well adapted to variable field conditions (Jaffe et al., 1990). However, as outlined above, many factors still have to be considered before introducing ants to control O. megnini populations in stabled horses and further studies are required.
Financial assistance from National Research Council (Grant 11-44) is gratefully acknowledged.
Adgaba N., Shenkute A.G., Al-Ghamdi A.A., Assefa A., Ansari M.J., Taylor B., Radloff S. 2014 — Crematogaster chiarinii ants as a potential biological control agent for protecting honeybee colonies from attack by Dorylus quadratus driver ants in Ethiopia (Hymenoptera: Formicidae) — Agric. For. Entomol., 16: 302-313. doi:10.1111/afe.12060 ![]()
Ariyarathne S., Apanaskevich D.A., Amarasinghe P.H., Rajakaruna R.S. 2016 — Diversity and distribution of tick species (Acari: Ixodidae) associated with human otoacariasis and socio-ecological risk factors of tick infestations in Sri Lanka — Exp. Appl. Acarol.: 1-25. doi:10.1007/s10493-016-0056-z ![]()
Barbosa P.A. 1998 — Agroecosystems and Conservation biological control — In: Barbosa P. (Eds). Conservation biological control Academic Press, London. doi:10.1016/B978-012078147-8/50049-9 ![]()
Bellows T.S., T.W. Fisher 1999 — Handbook of biological control: principles and applications of biological control — Academic, San Diego, CA.
Bishopp F.C. 1913 — The fowl tick (Argas miniatus Koch) — U.S. Dep. Agric. Bur. Entomol., 170: 1-14.
Bolton B. 1994 — Identification Guide to the Ant Genera of the World. Harvard University Press, USA.
Castineiras A., Jimeno G., Lopez M., Sosa L.M. 1987 — Effect of Beauveria bassiana, Metarhizium anisopliae (Fungi: Imperfecti) and Pheidole megacephala (Hymenopthera: Formicidae) on eggs of Boophilus microplus (Acarina: Ixodidae) — Rev. Salud Anim., 9: 288-293.
Dias R.K.S. 2002 — Ants of Sri Lanka with a brief description of field and laboratory methods — Sarvodaya Vishvalekha Press Ratmalana. pp. 44.
Diyes G.C.P., Karunaratne W.A.I.P., Tomberlin J.K., Rajakaruna R.S. 2015 — Case Study First record of Megaselia scalaris (LOEW)(DIPTERA: PHORIDAE) infesting a spinose ear tick, Otobius megnini, colony in Sri Lanka — Trop. Biomed., 32: 791-795.
Diyes G.C.P., Rajakaruna R.S. 2016a — Life cycle of Spinose ear tick, Otobius megnini (Acari: Argasidae) infesting the race horses in Nuwara Eliya, Sri Lanka — Acta Trop., 166:164-176. doi:10.1016/j.actatropica.2016.11.026 ![]()
Diyes G.C.P., Rajakaruna R.S. 2016b — Seasonal dynamics of Spinose ear tick Otobius megnini associated with horse otoacariasis in Sri Lanka — Acta Trop., 159: 170-175. doi:10.1016/j.actatropica.2016.03.025 ![]()
Estrada-Pena A., Jongejan F. 1999 — Ticks feeding on humans: a review of records on human-biting Ixodoidea with special reference to pathogen transmission — Exp. Appl. Acarol., 23: 685-715.
Fisher T.W., Bellows T.S., Caltagirone L.E., Dahlsten D.L., Huffaker, C.B., Gordh G. 1999 — Handbook of biological control: principles and applications of biological control — Academic Press, San Diego, California.
Foil L.D., Coleman P., Eisler, M., Fragoso-Sanchez H., Garcia-Vazquez Z., Guerrero F.D., Jonsson N.N., Langstaff I.G., Li A.Y., Machila N., Miller R.J., Morton J., Pruett J.H., Torr S. 2004 — Factors that influence the prevalence of acaricide resistance and tick-borne diseases — Vet. Parasitol., 125: 163-181.
Gleim E.R., Conner L.M., Yabsley M.J. 2013 — The effects of Solenopsis invicta (Hymenoptera: Formicidae) and burned habitat on the survival of Amblyomma americanum (Acari: Ixodidae) and Amblyomma maculatum (Acari: Ixodidae) — J. Med. Entomol., 50: 270-276. doi:10.1603/ME12168 ![]()
Gómez-Nuñez J.C. 1971 — Tapinoma melanocephalum as an inhibitor of Rhodnius prolixus populations — J. Med. Entomol., 8: 735-737.
Gromadzki T.Z., Bull C.M. 1997 — Ant predation on different life stages of two Australian ticks — Exp. Appl. Acarol., 21: 109-115.
Guerrero F.D., Lovis L., Martins J.R. 2012 — Acaricide resistance mechanisms in Rhipicephalus (Boophilus) microplus — Rev. Bras. Parasitol. Vet., 2: 1-6.
Holway D.A., Lach L., Suarez A.V., Tsutsui N.D., Case T.J. 2002 — The causes and consequences of ant invasions — Annu. Rev. Ecol. Syst.: 181-233. doi:10.1146/annurev.ecolsys.33.010802.150444 ![]()
Jaffe K., Mauleon H., Kermarrec A. 1990 — Qualitative evaluation of ants as biological control agents with special reference to predators on Diaprepes spp. (Coleoptera: Curculionidae) on citrus groves in Martinique and Guadeloupe. In Colloques de l'INRA; Caribbean Meetings on Biological Control., pp. 405-416.
Jellison W.L., Bell E.J., Huebner R.J., Parker R.R., Welsh H.H. 1948 — Q fever studies in Southern California. IV. Occurrence of Coxiella burneti in the spinose ear tick, Otobius megnini — Public Health Rep., 63: 1483-1489. doi:10.2307/4586756 ![]()
Jenkins D.W. 1964 — Pathogens, Parasites and Predators of Medically Important Arthropods. Annotated List and Bibliography — Bulletin of the WHO: 30.
Kaaya G.P. 2003 — Prospects for innovative tick control methods in Africa — Int. J. Trop. Insect Sci., 23: 59-67. doi:10.1017/S174275840001225X ![]()
Lovis L., Perret J.L., Bouvier J., Fellay J.M., Kaminsky R., Betscharta B., Sager H. 2011 — A new in vitro test to evaluate the resistance level against acaricides of the cattle tick, Rhipicephalus (Boophilus) microplus — Vet. Parasitol., 182: 269-280.
Madigan J.E., Valberg S.J., Ragle C., Moody J.L. 1995 — Muscle spasms associated with ear tick (Otobius megnini) infestations in five horses — J. Am. Vet. Med. Association., 207: 74-76.
Montañez M.L., Guerrero H.C., Luque Z., Méndez A. 1998. — Biological control of Leptopharsa gibbicarina (Hemiptera: Tingidae) with the ant Crematogaster sp. (Hymenoptera: Formicidae) in oil palm — Rev. Colomb. Entomol., 24: 89-94.
Mwangi E.N., Kaaya G.P. 1997 — Prospects of using tick parasitoids (Insecta) for tick management in Africa — Int. J. Acarol., 23: 215-219 doi:10.1080/01647959708683566 ![]()
Oliver J.H. Jr., McKeever S., Pound J.M. 1986 — Parasitism of larval Ixodes ticks by chigger mites and fed female Ornithodoros ticks by Ornithodoros males — J. Parasitol., 72: 811-812.
Parish H.E. 1949 — Recent studies on life history and habits of the ear tick — J. Econ. Entomol., 42: 416-419.
Rajput Z.I., Hu S., Chen W., Arijo A.G., Xiao C. 2006 — Importance of ticks and their chemical and immunological control in livestock — J. Zhejiang Uni. Sci., 7: 912-921.
Rodriguez J., Villalba G., Abrey R., Castaneiras A. 1983— Pheidole megacephala (Hymenoptera: Formicidae) as a predator of Amblyomma cajennense (Acarina: Ixodidae) in Cuba — Rev. Salud Anim. 5: 437-440.
Samish M., Alekseev E.A. 2001 — Arthropods as predators of ticks (Ixodoidea) — J. Med. Entomol., 38: 1-11.
Samish M., Ginsberg H., Glazer I. 2004 — Biological control of ticks — Parasitol. 129: 389-403.
Samish M., Rehacek J. 1999 — Pathogens and predators of ticks and their potential in biological control — Annu. Rev. Entomol., 44: 159-182. doi:10.1146/annurev.ento.44.1.159 ![]()
Shyma K.P., Gupta J.P., Singh V. 2015 — Breeding strategies for tick resistance in tropical cattle: a sustainable approach for tick control. J. Parasit. Dis. 39: 1-6. doi:10.1007/s12639-013-0294-5 ![]()
Sonenshine D.E. 1993 — Biology of ticks — New York Oxford: Oxford University Press.
Sutherst R.W., Maywald G. 2005 — A climate model of the red imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae): implications for invasion of new regions, particularly Oceania — Environ. Entomol., 34: 317-335.
Symondson W.O.C., Sunderland K.D., Greenstone M.H. 2002 — Can generalist predators be effective biocontrol agents? — Annu. Rev. Entomol., 47: 561-594.
Wilkinson P.R. 1957 — The spelling of pasture in cattle tick control — Crop and Pasture Sci. 8: 414-423. doi:10.1071/AR9570414 ![]()
Wilkinson P.R. 1970 — Factors affecting the distribution and abundance of the cattle tick in Australia: observations and hypotheses — Acarol. 12: 492-507
Willadsen P., Bird P., Cobon G.S., Hungerford J. 1995 — Commercialisation of a recombinant vaccine against Boophilus microplus — Parasitol. 110 Suppl:S43-50
Zaman M.A., Iqbal, Z., Abbas R.Z., Khan M.N., Muhammad G., Younus M., Ahmed S. 2012 — In vitro and in vivo acaricidal activity of a herbal extract — Vet. Parasitol. 186: 431-436.



2016-08-28
Date accepted:
2017-02-11
Date published:
2017-07-06
Edited by:
McCoy, Karen

This work is licensed under a Creative Commons Attribution 4.0 International License
2017 Diyes, G.C.P.; Karunarathna, N.B.; Silva, T.H.S.E.; Karunaratne, W.A.I.P. and Rajakaruna, R.S.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)