




Patterns of soil mite diversity in Lamto savannah (Côte d’Ivoire) submitted at different fire regimes
N'Dri, Julien K.1 ; N'Da, Rodolphe Arnaud G.2 ; Seka, Fabrice A.3 ; Pokou, Pacôme K.4 ; Tondoh, Jérôme E.5 ; Lagerlöf, Jan6 ; Kone, Mouhamadou7 ; Dosso, Kanvaly8 ; N'Dri, Brigitte A.9 and Kone, N'Golo A.10
1✉ UFR des Sciences de la Nature Université Nangui Abrogoua 02 BP 801 Abidjan 02 Côte d’Ivoire.
2UFR des Sciences de la Nature Université Nangui Abrogoua 02 BP 801 Abidjan 02 Côte d’Ivoire.
3UFR des Sciences de la Nature Université Nangui Abrogoua 02 BP 801 Abidjan 02 Côte d’Ivoire.
4UFR des Sciences de la Nature Université Nangui Abrogoua 02 BP 801 Abidjan 02 Côte d’Ivoire.
5UFR des Sciences de la Nature Université Nangui Abrogoua 02 BP 801 Abidjan 02 Côte d’Ivoire.
6Swedish University of Agricultural Sciences (SLU), Dept. of Ecology, P.O.Box 7044, SE-750 07 Uppsala, Sweden.
7Université Peleforo Gon Coulibaly, Côte d’Ivoire.
8UFR des Sciences de la Nature Université Nangui Abrogoua 02 BP 801 Abidjan 02 Côte d’Ivoire.
9UFR des Sciences de la Nature Université Nangui Abrogoua 02 BP 801 Abidjan 02 Côte d’Ivoire.
10UFR des Sciences de la Nature Université Nangui Abrogoua 02 BP 801 Abidjan 02 Côte d’Ivoire.
2017 - Volume: 57 Issue: 4 pages: 823-833
https://doi.org/10.24349/acarologia/20174196Keywords
Abstract
Generally fire is seen as an important management tool and biodiversity conservation (Driscoll et al., 2010; Hugo-Coetzee and Avenant, 2011). Its use and its various applications favor landscape heterogeneity, namely diversification of vegetation and soil biodiversity (Hudak et al., 2004). In addition to their impact on vegetation, fires affect different soil horizons. Indeed, they modify the physico-chemical and biological properties of soil (Allison et al., 2010; Certini et al., 2011; Chen and Shrestha, 2012; Comte et al., 2012), notably by the alteration of the quantity and quality of organic matter, and carbon stock. Whether applied in a herbaceous habitat (Hugo-Coetzee and Avenant, 2011) or in natural forest (Malmstrom et al., 2009; Malmstrom, 2010, 2012), a better understanding of species response to different fire regimes is essential for sustainable environmental management (Driscoll et al., 2010). Several studies have shown the reduction in the abundance and diversity of soil mites after the passage of fire in different vegetation (Malmstrom et al., 2009; Grabczynska et al., 2009; Michael et al., 2012).
In Côte d’Ivoire, Lamto savannah is one of the most protected areas. The management of this ecosystem includes the use of fire. Indeed the fire is artificially triggered and controlled towards mid January. This type of fire is locally called « mid season fire » and generally applied to relatively dry vegetation. The mid season fire destroyed much of the herbaceous and stratum. However, it must be noted that regularly burned patches tend to become afforested. In less than thirty years, unburned plots to Lamto were colonized by forest species (Vuattoux et al., 2006). This environmental change would be partly due to climate change (Le Roux, 2006). In this context, could the change in fire regime promote the maintenance of Lamto savannah? Except for Collins et al. (2012), very few studies make references to the change of fire regime. Nonetheless the variation in fire severity could explain the species variation observed or rediscovered after the passage of fire (Malmstrom, 2010). Unfortunately this factor is not taken into account in many studies concerning the impact of fire on soil organisms. Soil life is intimately linked to living organisms that play a major role (Gulvik, 2007; Gergocs and Hufnagel, 2009). An analysis of their response to fire regime change will promote the savannah environmental monitoring. Besides the « mid season fire », this investigation aims to test the application of two other types of fire that are the « early fire » and the « late fire » respectively established in November and March. The late fire is fire applied to highly dry vegetation whereas the early fire is fire applied to humid vegetation with a green appearance.
The main objective of this investigation was to evaluate the impact of different fire regimes on soil mite abundance and diversity. Two hypotheses underlie this study (i) the passage of a mid season fire regime with an early or late fire regime causes a variation in the abundance and diversity of soil mites (ii) whatever the fire regime soil mite abundance and diversity decrease after the passage of fire.
With an area of 2,500 ha, the Lamto reserve is located at 160 km northwest of Abidjan (5°02 W, 6°13 N), in the extreme south of "V Baoulé" (Advanced area of the savannah in the rainforest). The climate is humid and intertropical (Le Roux, 2006). The monthly rainfall ranged from 27.32 in December to 189.4 mm in June during the last ten years preceding field work. The average monthly rainfall was about 104.79 mm over last decade. During the study year (2013-2014) the monthly rainfall varied between 22.65 mm in December to 197.80 mm in May with an average monthly of 87.40 mm. Average monthly temperature (2013-2014) was about 29°C. The vegetation of Lamto is a forest-savannah mosaic (Menaut and César, 1979), (i) the gallery forests (ii) herbaceous savannah dominated by Loudetia simplex (iii) shrub savannahs dominated by Hyparrhenia diplanda and Andropogon sp. (iv) wooded savannah (v) shrub savannahs protected from fire. Soils are ferralitic type under forests and tropical ferruginous type under savannah (Delmas, 1967; Riou, 1974). The sampling was conducted in shrub savannah of Lamto not protected from fire.
In this area, three sites (Salty marigot, Plateau and North piste) were selected and used as replicates. The choice of these sites allowed taking into account environmental heterogeneity. The site of Plateau was more wooded whereas Salty marigot and North piste were more grassy. On each site, three adjacent plots of 100 m x 50 m were delimited for the sampling. A layer of 10 m width was made around each plot. Five sampling point were identified on each plot following the two diagonals. The three fire regimes were applied respectively to three plots and three sites. Early fire was applied in November, the mid season fire in January and late fire in March. At each fire regime, soil cores were taken three days before the fire application, the day after the fire and one month after the passage of fire. At each sampling point, two adjacent soil cores (1m interval) were taken with a steel corer (Ø 4 cm) at 0-10 cm soil depth, hence a total of 10 soil cores per plot. Thus 10 soil cores x 3 sampling periods x 3 fire regimes or plots x 3 sites gave a total of 270 soil cores. These soil cores were for mite extraction. Three days before fire application, three soil cores were taken following two upper layers (0-5 and 5-10 cm) on the first diagonal by using the cylinder method (Assié et al., 2008) for physical measurements. These same samples were performed on the opposite diagonal one month after the passage of fire. 108 soil cores were sampled for estimation of the bulk density and water content. Fire intensity data were provided by Soro (2015).
Soil mites were extracted by using modified Berlese-Tullgren funnel over 10 days (Bedano and Ruf, 2007; N’Dri et al., 2011a) and stored in a solution of 70% alcohol. All mites from samples were mounted on cavity slides containing 85% lactic acid solution. Oribatid mites, darker and more robust species were cleared in 85% lactic acid solution for 24-72 hours, depending upon the level of sclerotization of each species and observed with a digital camera VC.5000 mounted on a NOVEX light microscope. SEM pictures were made at the Royal Museum for Central Africa, Belgium. Only adult mites were taken in account during the identification. In the absence of African keys, adult mites were identified to major groups (Actinedida, Gamasida, Oribatida and Acaridida), family, genus and morphospecies levels by using keys and illustrations provided in Balogh and Balogh (1992), Krantz (1978) and Walter et al. (2013). Major groups (Oribatida, Acaridida) were defined in the traditional sense used in soil zoology and available in Krantz (1978) and Dindal (1990), not in the more recent sense found in Krantz and Walter (2009).
Along the three fire regime, soil mite abundance was estimated by using density expressed as mean individuals per square meter. Mite diversity was assessed by using Mean species richness, Cumulated species richness, Simpson index, Simpson diversity index and Evenness. Abundance data were normalized following the function ln (x + 1) after verification of homogeneity test (Levene test). Impact of fire regimes and sampling periods on soil mite abundance were evaluated by using a one-way ANOVA associated with the post-hoc Fisher test and Least Significant Difference test. The same analysis was used to compare diversity variables and physical parameters. Bulk density and water content were evaluated by depth (0-5 and 5-10cm) using the Student t-test. All tests were conducted using the software Statistica 7.1. (StatSoft Inc., Tulsa, USA). The software EstimateS 7.5 (Colwell, 2005) was used to estimate Cumulated species richness (Sob) in all soil cores from each fire regime after 500 randomization.
Soil mites mean density varied (Anova 1; F = 3.93, P = 0.020) significantly through the different fire regimes. The highest mean density of soil mites was recorded during the mid season fire (1141 ± 169 ind.m-2) whereas the lowest value was observed through the late fire (513 ± 86 ind.m-2). Whatever the fire regime, mite densities varied significantly (Anova 1; early fire: F = 23.78, P = 0.0000001; mid season fire: F = 3.38, P = 0.038; late fire: F = 5.52, P = 0.005) between the sampling period (Figure 1). In all cases highest densities were recorded before the passage of the fire (early fire: 1618 ± 176 ind.m-2; mid season fire: 1778 ± 316 ind.m-2; late fire: 769 ± 180 ind.m-2).

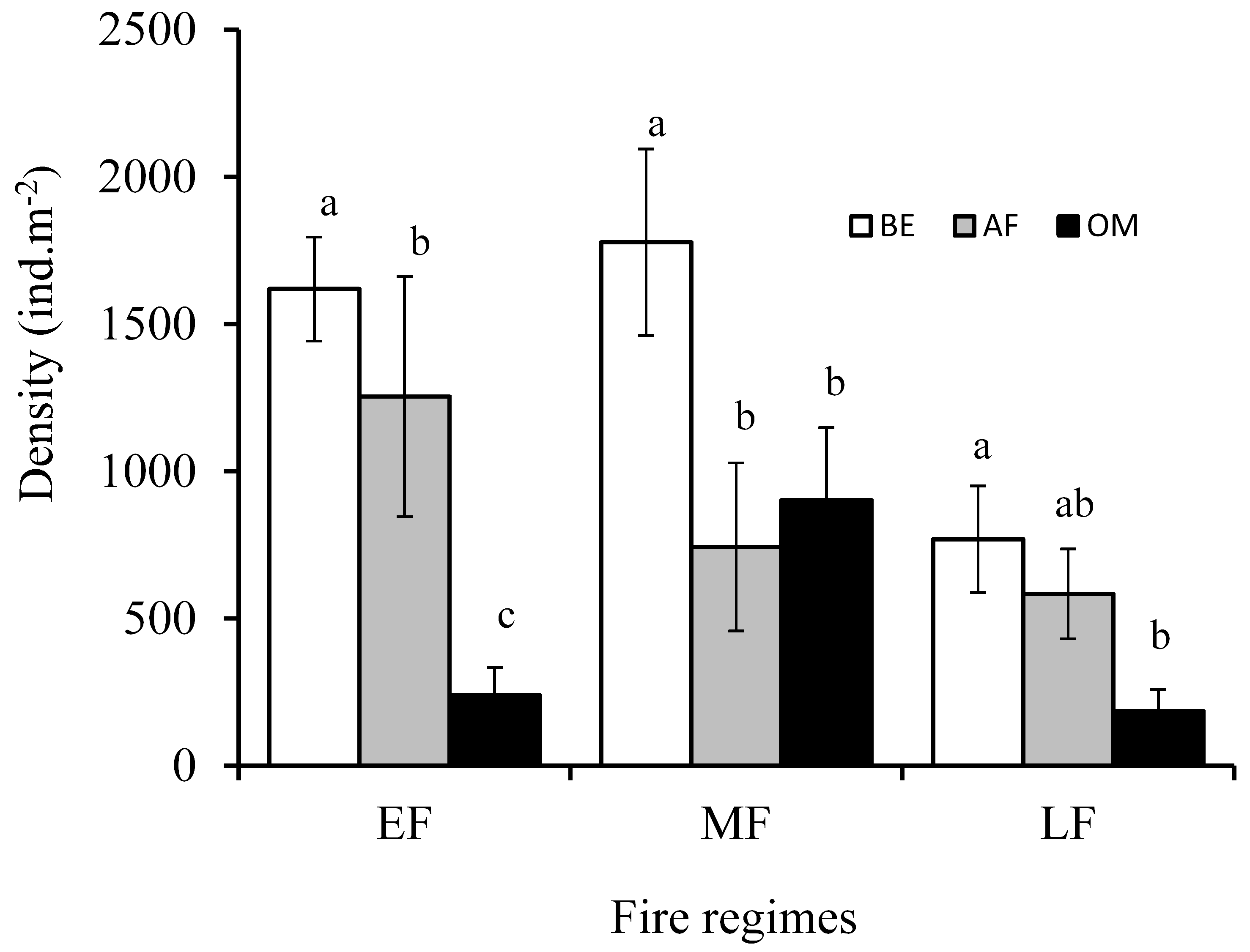
Mean species richness differed significantly (Anova 1; F = 4.71, P = 0.009) across the fire regimes and varied on the three sampling periods (Anova 1; early fire: F = 10.74, P = 0.000068; mid season fire: F = 4.80, P = 0.010; late fire: F = 4.37, P = 0.015). The greater mean richness (Table 1) were observed before the passage of fire (early fire: 1.86 ± 0.20 species; mid season fire: 1.9 ± 0.36 species; late fire: 0.9 ± 0.2 species). Except for mid season fire (Anova 1; F = 0.44, P = 0.639), Simpson index values were substantially modified across the three sampling periods (Anova 1; early fire: F = 8.29, P = 0.0005; late fire: F = 3.20, P = 0.045). However the reverse trend was observed (Anova 1; F = 2.79, P = 0.062) if we considered the three fire regimes. A part from late fire (Anova 1; F = 2.45, P = 0.09), Evenness values were modified significantly along the three sampling periods (Anova 1; early fire: F = 12.89, P = 0.000012; mid season fire: F = 5.10, P = 0.0079) and the three fire regimes (Anova 1; F = 3.34, P = 0.036).


Fire impacted differently the diversity parameters of major groups. Only mean species richness (Anova 1; F = 5.94, P = 0.0038), Simpson diversity index (Anova 1; F = 5.16, P = 0.0075) and Evenness (Anova 1; F = 3.81, P = 0.0257) from Gamasida were significantly modified along the three fire regime (Table 2). Gamasida and Oribatida were the most diversified groups. Mean species richness (0.26 ± 0.09 to 1.06 ± 0.21) and Simpson diversity index (0.01 ± 0.01 to 0.18 ± 0.05) of Gamasida ranged respectively from late fire to early fire. Highest values of mean species richness (2.46 ± 0.46) and Simpson diversity index (0.32 ± 0.06) from Oribatida were observed during the mid season fire whereas lowest values of mean species richness (1.33 ± 0.28) and Simpson diversity index (0.19 ± 0.05) were recorded during the late fire.


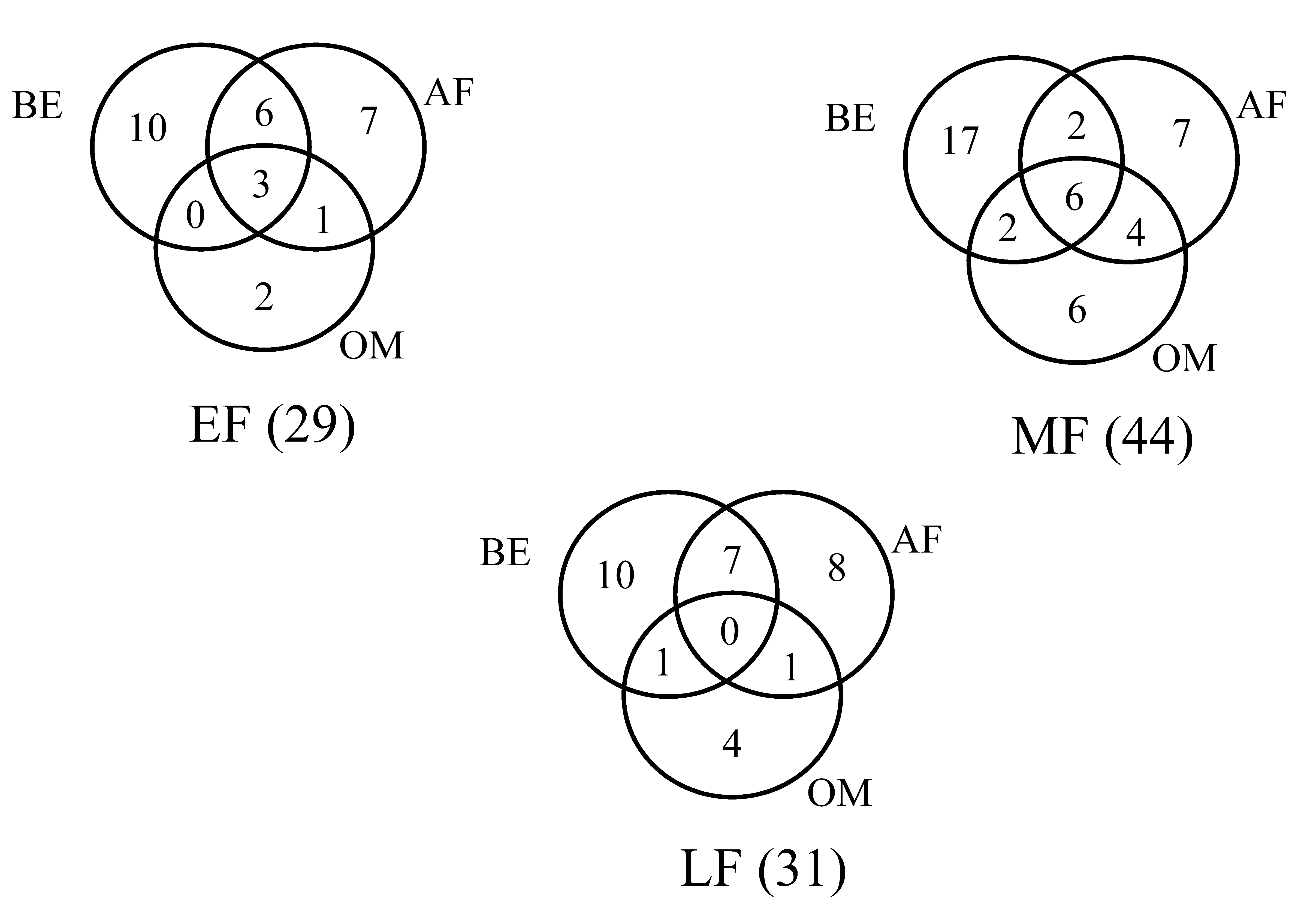
Whatever the fire regimes, the Oribatida and Gamasida represented the dominant groups (early fire: Gamasida 35%, Oribatida 55%, mid season fire: Gamasida 16%, Oribatida 70%, late fire: Gamasida 16%, Oribatida 74%). 70 morphospecies were identified across the three fire regimes (see appendix). 29, 44 and 31 species were observed respectively during the early, mid season and late fires. The lower Oribatida represented 25% of the total Oribatida. 100% of higher Oribatida was recorded the day after the fire and one month after the passage of late fire.
Oribatida was consisted of five groups (Palaeosomata, Parhyposomata, Desmonomata, Brachypylina and Poronota). Whatever the fire regimes, brachypyline adult abundance increased the day after the fire application. Respectively three and six common species were observed during the early and mid season fires (Figure 2).

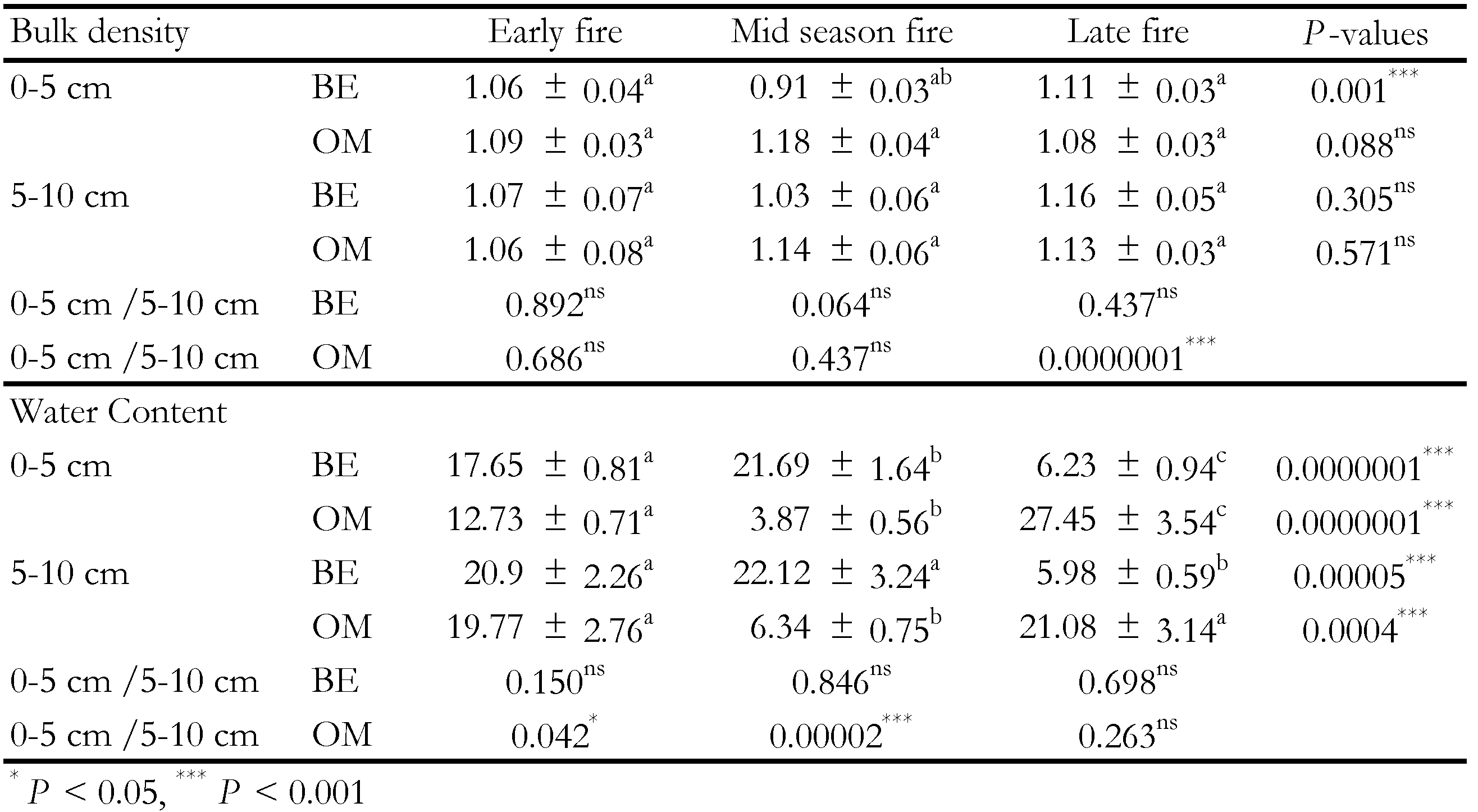
Bulk density values were substantially identical in the two layer sampled and varied from 0.91 ± 0.03 to 1.18 ± 0.04 g.cm-3 respectively before and one month after the mid season fire. Water content values were highest (27.45 ± 3.54%) and lowest (3.87 ± 0.56%) respectively one month after the early and mid season fire (Table 3). The physical variables analysis revealed that the bulk density was significantly (Anova 1; F = 9.29, P = 0.001) modified before the fire application on the upper layer (0-5 cm). One month after the late fire, we noticed a significant (t = -96.62, P = 0.0000001) increase of bulk density values with soil depth. Whatever the sampling period and soil layers, water content values changed significantly across the three fire regimes. As with the bulk density, soil water content values increased significantly with soil depth one month after (early fire: t = -1.58, P = 0.042; mid season fire: t = -8.88, P = 0.00002).

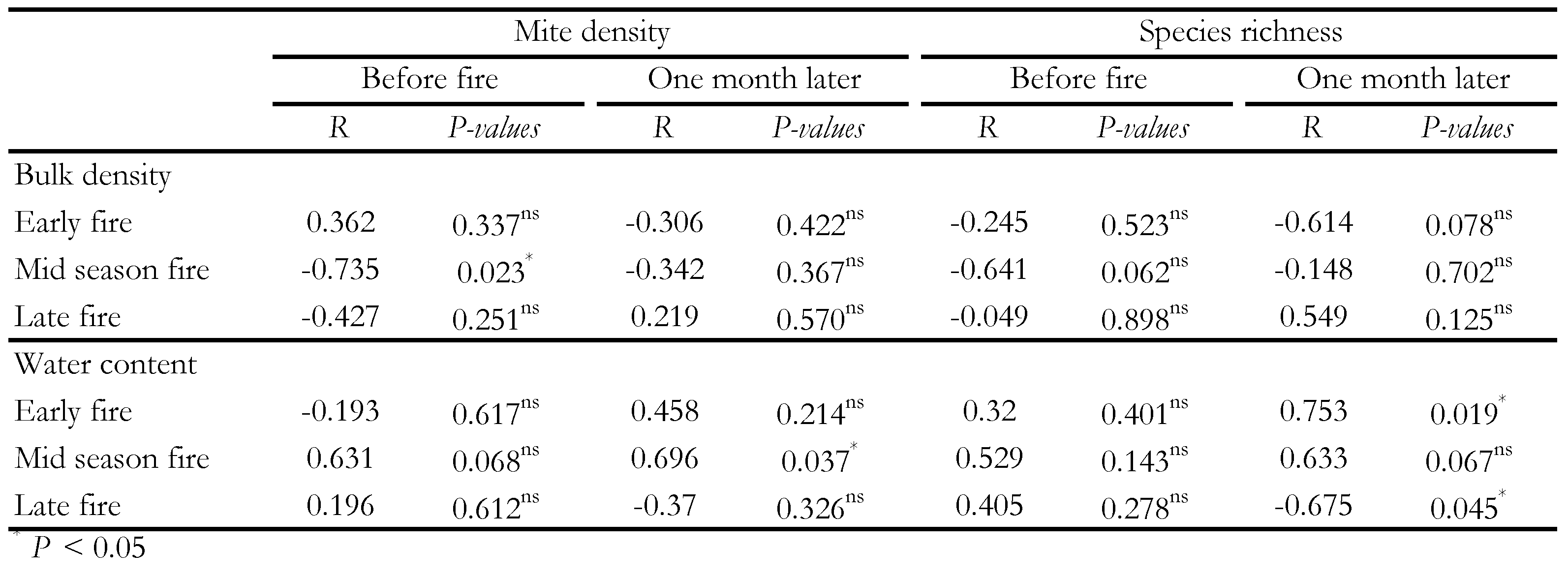
The different interactions showed the existence of a significant correlation between fire intensity and mite density and species richness respectively after the application of the early fire and one month after the passage of the late fire (Table 4). Soil mite density was significantly influenced by the bulk density and water content respectively before and one month after the mid season fire application (Table 5). However, the species richness was governed only by the water content, respectively one month after the early and late fires.



The evaluation of various soil mite communities revealed significant modification in mite density and richness (Grabczynska et al., 2009; Michael et al., 2012) after fire application. The result of this investigation indicated that mite density and richness decrease significantly after the fire application. These observations were similar with the results obtained by Malmstrom et al. (2009), Grabczynska et al. (2009) and Michael et al. (2012). According to these authors, the fire was seen as a factor altering the mite species. Indeed from a high intensity fire result a low abundance of soil mites. The response of soil microarthropods to the fire application (disturbance) could vary according the species is soft or sclerotized (Parisi et al., 2005). The eco-morphologic index (EMI) of edaphic microarthropod differed according the species are eu-edaphic (i.e. deep soil-living forms), hemi-edaphic (i.e. intermediate forms) or epi-edaphic (surface-living forms) (Parisi et al., 2005). From this point of view, soil temperature could be seen as a factor controlling the development of mite species (Athias, 1975, 1976; Oke et al., 2005) since the tolerable limit in the Lamto savannah was 39 °C with a pF between 2.4 and 4.2. These arguments agree with investigation made by Malmström (2008) who had used the forest humus at different temperatures in order to determine the lethal temperatures of microarthropods. The works of Ermilov and Łochynska (2008) had helped to highlight the influence of temperature on the Oribatida species development from the egg stage to adult. Thereby at 20°C the life cycle of species from genus Ceratoppia was shorter than those of Nanhermannia coronata (Berlese, 1913). The high intensity fire (4269 Kw.min-1) consecutive to drier vegetation could explain the low density observed during the late fire.
The highest density was recorded during the mid season fire. Traditionally, the mid season fire was practiced in Lamto savannah each year for over 30 years. Probably the great abundance of sclerotized mites from brachypiline and poronota groups could explain the highest density observed during the mid season fire. This argument was supported by the lack of significant correlation between the fire intensity and the mite density and species richness observed. Nonetheless, the densities observed before the fire application were inferior to those from the works performed by Noti et al. (1996) in a humid savannah of the Democratic Republic of Congo (dry season: 7500 ind.m-2; rainy season: 13500 ind.m-2). 70 species had been recorded along the three fire regimes and distributed as follows: early fire 29 species vs. mid season fire 44 species vs. late fire 31 species. The significant difference of species richness observed along the fire regime indicated a heterogeneous response of soil mite to fire regime change. Overall the observed species richness remained inferior to those recorded by Noti et al. (2003) in the same humid savannah (105 Oribatida species). The fundamental reason could be the poverty of Lamto soil in nutrients (Riou, 1974; Mordelet et al., 1996) consecutive to annual fire application. Indeed the fire destroys the amount and quality of the available soil organic matter (Certini et al., 2011; Chen and Shrestha, 2012). While this chemical component remains an essential support for the maintenance and development of Oribatida species (Behan-Pelletier, 1999). The results also revealed that soil mites were strongly dependent on the bulk density and water content. These trends were similar to observations made by several authors (Noti et al., 2003; N’Dri et al., 2011a). During the late fire, soils seem less porous (bulk density, 0-5 cm: 1.11 g.cm-3 vs. 5-10 cm: 1.16 g.cm-3) and less humid (water content, 0-5 cm: 6.23% vs. 5-10 cm: 5.98%). A comparison of recent work provided by N’Dri et al. (2011a,b) in shrub savannah protected from fire and during the same season revealed a more porous soil (bulk density, 0-5 cm: 0.70 g.cm-3 vs. 5-10 cm: 0.93 g.cm-3) relatively more wet (water content, 0-5 cm: 9.80% vs. 5-10 cm: 8.80%) with 85 species recorded. The heterogeneous distribution of species numbers reflects an alteration of soil porosity and organic matter (Riou, 1974; Mordelet et al., 1996).
Fire intensity reduced drastically soil mite abundance and diversity. However its impact was influenced by the structure and quality of soil and the edaphic microclimate. Nonetheless an estimation of soil litter in time, followed by an evaluation of vegetation cover, particularly the regrowth rate after the fire application could better inform us on the appropriate type of fire about the management of the Lamto savannah.
Our sincere thanks to Pr Yéo extscKolo, Director of the Lamto Ecology Station who kindly allowed access to the different study site. A Big thanks to Drs. extscMartinez A. extscGuéi and Ettien F. extscEdoukou
Allison S.D., McGuire K.L., Treseder K.K. 2010 — Resistance of microbial and soil properties to warming treatment seven years after boreal fire — Soil Biol. Biochem., 42: 1872-1878. doi:10.1016/j.soilbio.2010.07.011 ![]()
Assié K.H., Angui P., Tamia A.J. 2008 — Effets de la mise en culture et des contraintes naturelles sur quelques propriétés physiques d'un sol ferrallitique au Centre Ouest de la Côte d'Ivoire: Conséquences sur la dégradation des sols — Eur. J. Sci. Res., 23: 149-166.
Athias F., Josens G., Lavelle P. 1975 — Traits généraux du peuplement animal endogé de la savane de Lamto (Côte d'Ivoire). In Vanek J. (Ed.). Progress in Soil Zoology — Academia Publishing House, Prague, Czech Republic, pp. 375-388. doi:10.1007/978-94-010-1933-0_42 ![]()
Athias F. 1976 — Recherche sur les microarthropodes du sol de la savane de Lamto (Côte- d'Ivoire) — Ann. Univ. Abidjan, sér. E, écologie, 9: 193-271.
Balogh J., Balogh P. 1992 — The Oribatid Mites Genera of the World (vol. 1 and 2) — The Hungarian National Museum Press, Budapest, Hungary.
Bedano J.C., Ruf A. 2007 — Soil predatory mite communities (Acari: Gamasina) in agroecosystems of Central Argentina — Appl. soil ecol., 36: 22-31. doi:10.1016/j.apsoil.2006.11.008 ![]()
Behan-Pelletier V.M. 1999 — Oribatid mite biodiversity in agroecosystems: role for bioindication — Agr. Ecosyst. Environ., 74: 411-423. doi:10.1016/S0167-8809(99)00046-8 ![]()
Certini G., Nocentini C., Knicker H., Arfaioli P., Rumpel C. 2011 — Wildfire effects on soil organic matter quantity and quality in two fire-prone Mediterranean pine forests — Geoderma, 167-168: 148-155.
Chen H.Y.H., Shrestha B.M. 2012 — Stand age, fire and clearcutting affect soil organic carbon and aggregation of mineral soils in boreal forests — Soil Biol. Biochem., 50: 149-157. doi:10.1016/j.soilbio.2012.03.014 ![]()
Collins L., Bradstock R.A., Tasker E.M., Whelan R.J. 2012 — Impact of fire regimes, logging and topography on hollows in fallen logs in eucalypt forest of south eastern Australia — Biol. Conserv., 149: 23-31. doi:10.1016/j.biocon.2012.01.065 ![]()
Colwell R.K. 2005 — EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. Version 7.5 — Persistent URL: https://purl.oclc.org/estimates ![]() .
.
Comte I., Davidson R., Lucottea M., Reis de Carvalhod C.J. 2012 — Physicochemical properties of soils in the Brazilian Amazon following fire-free land preparation and slash-and-burn practices — Agr. Ecosyst. Environ., 156: 108-115. doi:10.1016/j.agee.2012.05.004 ![]()
Delmas J. 1967 — Recherches écologiques dans la savane de Lamto: premier aperçu sur les sols et leur valeur agronomique — La Terre et la Vie, 3: 216-227.
Dindal D.L. (ed) 1990 — Soil Biology Guide — Wiley J. & Sons, New York, U.S.A.
Driscoll D.A., Lindenmayer D.B., Bennett A.F., Bode M., Bradstock R.A., Cary G.J., Clarke M.F., Dexter N., Fensham R., Friend G. 2010 — Fire management for biodiversity conservation: Key research questions and our capacity to answer them — Biol. Conserv., 143: 1928-1939.
Ermilov S.G., Łochynska M. 2008 — The influence temperature on the development time of three Oribatid mite species (Acari, Oribatida) — North-western J. Zool., 2: 274-281.
Gergócs V., Hufnagel L. 2009 — Application of oribatid mites as indicators (review) — Appl. Ecol. Environ. Res., 7(1): 79-98. doi:10.15666/aeer/0701_079098 ![]()
Grabczynska O., Oleniczak I., Predecka A., Russel S. 2009 — Short-term effects of prescribed forest fire on soil mite (Acari) — Short Res. Contribution, 57(4): 805-809.
Gulvik M.E. 2007 — Mites (Acari) as indicators of soil biodiversity and land use monitoring: A review — Pol. J. Ecol., 55(3): 415-440.
Hudak A.T., Fairbanks D.H.K., Brockett B.H. 2004 — Trends in fire patterns in a southern African savanna under alternative land use practices — Agr. Ecosyst. Environ., 101: 307-325. doi:10.1016/j.agee.2003.09.010 ![]()
Hugo-Coetzee E.A., Avenant N.L. 2011 — The effect of fire on soil oribatid mites (Acari: Oribatida) in a South African grassland — Zoosymposia, 6: 210-220.
Krantz G.W. 1978 — A Manual of Acarology (2nd ed.) — Oregon State University Bookstores, Corvallis, U.S.A.
Le Roux X. 2006 — Climate. In Abbadie L., Gignoux J., Le Roux X., Lepage M. (Eds). Lamto. Structure, Functioning, and Dynamics of a Savanna Ecosystem — Springer Verlag, New York, USA, pp. 25-44.
Malmström A. 2008 — Temperature tolerance in soil microarthropods: Simulation of forest -fire heating in the laboratory — Pedobiologia, 51: 419-426.
Malmström A., Persson T., Ahlström K., Gongalsky K.B., Bengtsson J. 2009 — Dynamics of soil meso- and macrofauna during a 5-year period after clear-cut burning in a boreal forest — Appl. Soil Ecol., 43: 61-74. doi:10.1016/j.apsoil.2009.06.002 ![]()
Malmström A. 2010 — The importance of measuring fire severity-Evidence from microarthropod studies — forest Ecol. Manage., 260: 62-70.
Malmström A. 2012 — Life-history traits predict recovery patterns in Collembola species after fire: A 10 year study — Appl. Soil Ecol., 56: 35-42. doi:10.1016/j.apsoil.2012.02.007 ![]()
Menaut J.C., César J. 1979 — Structure and primary productivity of Lamto savannas, Ivory Coast — Ecology, 60(6): 1197-1210.
Michael A.C., Lamoncha K.L., Gillette N.E. 2012 — Oribatid Mite Community Decline Two Years after Low-Intensity Burning in the Southern Cascade Range of California, USA — Forests, 3: 959-985.
Mordelet P., Barot S., Abbadie L. 1996 — Root foraging strategies and soil patchiness in a humid savanna — Plant Soil, 182: 171-176. doi:10.1007/BF00011005 ![]()
N'Dri J.K. 2010 — Diversité biologique des acariens du sol de la Côte d'Ivoire — Mémoire de Thèse Unique de doctorat de la faculté des Sciences. École Doctorale en Sciences, Département de Biologie, Earth and Life Institut, Centre de Recherche sur la Biodiversité. Université Catholique de Louvain, Belgique, n° 214, 177p.
N'Dri J.K., Andre H.M. 2011a — Soil mite densities from central Ivory Coast — J. Anim. Plant Sci., 10(2): 1283-1299.
N'Dri J.K., Andre H.M., Hance T. 2011b — Soil mite diversity from Ivory Coast — Eur. J. Scientific Res., 64(2): 263-276.
Noti M-L., André H.M., Dufrêne M. 1996 — Soil oribatid mite communities (Acari: Oribatida) from high Shaba (Zaire) in relation to vegetation — Appl. Soil Ecol., 5: 81-96. doi:10.1016/S0929-1393(96)00122-9 ![]()
Noti M-I., André H.M., Ducarme X., Lebrun Ph. 2003 — Diversity of soil oribatid mites (Acari: Oribatida) from high Katanga (Democratic Republic of Congo): a multiscale and multifactor approach — Biodiv. Conserv., 12: 767-785.
Oke O.A., Adejuyigbe C.O., Ajede O.A. 2005 — Survey of soil micro-arthropods in selected sites in ogun state — Paper prepared for presentation at the Farm Management Association of Nigeria Conference, Asaba, Nigeria, October 18-20, pp.12-19.
Parisi V.C., Menta C., Gardi C., Jacomini C., Mozzanica E. 2005 — Microarthropod communities as a tool to assess soil quality and biodiversity: a new approach in Italy — Agr. Ecosyst. Environ., 105: 323-333. doi:10.1016/j.agee.2004.02.002 ![]()
Riou G. 1974 — Les sols de la savane de Lamto — Bulletin de Liaison des Chercheurs de Lamto, 1: 3-45.
Soro D. 2015 — Caractéristiques du combustible et comportement du feu en fonction de différents régimes de feu dans une savane humide (Lamto, Côte d'Ivoire) — Mémoire de master1, Université Nangui Abrogoua, Abidjan, Côte d'Ivoire.
Vuattoux R., Konaté S., Abbadie L., Barot S., Gignoux J., Lahoreau G. 2006 — History of the Lamto Ecology Station and Ecological Studies at Lamto. In Abbadie L., Gignoux J., Le Roux X., Lepage M. (Eds). Lamto. Structure, Functioning, and Dynamics of a Savanna Ecosystem — Springer Verlag, New York, USA, pp. 1-9. doi:10.1007/978-0-387-33857-6_1 ![]()
Walter D.E., Latonas S., Byers K. 2013 — Almanac of Alberta Oribatida Part1. Ver. 2.3 — The Royal Aberta Museum, Edmonton, AB. ( http://www.royalabertamuseum.ca/natura/insects/reasearch/reasearch.htm ![]() )
)



2016-11-16
Date accepted:
2017-03-20
Date published:
2017-07-04
Edited by:
Magalhães, Sara

This work is licensed under a Creative Commons Attribution 4.0 International License
2017 N'Dri, Julien K.; N'Da, Rodolphe Arnaud G.; Seka, Fabrice A.; Pokou, Pacôme K.; Tondoh, Jérôme E.; Lagerlöf, Jan; Kone, Mouhamadou; Dosso, Kanvaly; N'Dri, Brigitte A. and Kone, N'Golo A.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)